


Abstract
Objectives: To optimize an enzyme-linked immunosorbent assay (ELISA) for measuring the HspX protein (α-crystallin) levels and then evaluate its correlation with the accumulation of lipid bodies in Mycobacterium bovis (M. bovis) during hypoxia and exposure to nitric oxide.
Methods: This study was conducted at Prince Sultan Military Medical City, Riyadh, Saudi Arabia between 2016 and 2017. We first optimized ELISA conditions for the detection of HspX. The optimization protocol focused on minimizing concentrations of the capture antibody, detection antibody, and conjugated secondary antibody, and determining the minimum detection limit of the antigen, HspX. Bacteria were grown either in shaking culture or in stationary flasks mimicking hypoxic environments. A standard Bradford assay was used to determine the total protein and HspX was detected using the optimized ELISA protocol. The effect of hypoxic environment and nitric oxide on the levels of HspX and lipid bodies, detected by staining with Nile red, was also evaluated.
Results: An optimized ELISA protocol was established for the detection of HspX from M. bovis. Exposure to nitric oxide and hypoxic conditions led to an increase in the levels of HspX protein. The increase in HspX associated with nitric oxide treatment and hypoxic conditions correlated with higher levels of lipid bodies mainly found in pathogenic mycobacteria.
Conclusions: The optimized ELISA protocol in this study can detect HspX protein levels in M. bovis growing in normal and hypoxic environments. Importantly, hypoxia led to enhanced expression of HspX protein, which correlated with the enhanced production of lipid bodies. Lipid body production is a survival strategy of pathogenic mycobacteria.
Mycobacterium tuberculosis (Mtb) is an obligate pathogen that can infect diverse human body sites, especially the lungs.1 Like Mtb, Mycobacterium bovis and Mycobacterium africanus can also cause tuberculosis (TB) in humans.2 Mycobacteria are the most dangerous bacterial pathogens, killing more people annually than any other infectious disease. It has been previously estimated that Mtb infects approximately one-third of the global population, and 10% of infected patients where the bacteria remain in the latent and not infectious phase will go on to develop active TB.3 However, with effective management, Mtb infection is being curtailed, and in 2016 it was estimated that one-quarter of the world’s population continues to suffer from Mtb infection.4
The Mycobacteriaceae family consists of more than 170 recognized species.5 Among these, a group of bacteria pathogenic in humans and other animals called the Mycobacterium tuberculosis complex (MtbC) contains at least 9 species: M. mungi, M. microti, M. pinnipedii, M. bovis, M. canetti, M. africanum, M. tuberculosis sensu stricto, M. caprae, and M. orygis.6 The MtbC members have the same 16s rRNA sequence but vary pathogenetically and phenotypically.7
Mycobacterium tuberculosis colonizes the human lower respiratory tract. The mechanism of infection begins with the inhalation of droplet containing the bacteria. Once the host inhales these droplets, they reach the pulmonary alveoli. An alveolar macrophage then ingests the bacterium, and it can survive without growth within the macrophage.8 This provides the pathogenic bacterium with a shield from the host immune system. The pathogen remains in this latent phase in healthy individuals until the host becomes immunocompromised, resulting in the organism’s activation and appearance (secondary infection).9
An infectious microorganism, including bacteria, is challenged by the body’s immune system and internal environment. For their survival, bacteria have evolved to produce specific biochemical changes in their cellular system in response to stress or host responses. Among these, excessive production of lipids is considered as the most effective strategy. It has been reported to play an essential role in developing resistance in pathogens like mycobacteria and several other species.10 The accumulation of lipid bodies in pathogenic mycobacteria and factors triggering this is an active area of investigation.
The HspX protein (also known as α-crystallin, Acr1) is predominantly found in mycobacteria and is involved in the survival of mycobacteria.
Our study aimed to develop most reliable and easy-to-use detection method for HspX and the association of HspX with the accumulation of lipid bodies involved in the survival strategies of these pathogens. We also investigated the production of lipid bodies necessary for mycobacterial survival.
Methods
Various experiments performed in this study were conducted at the Prince Sultan Military Medical City, Riyadh, KSA between 2016 and 2017. The study design is prospective and analytical. The Acr1 antibody catalog no. orb334091 from Biorbyt, UK; Goat anti-mouse IgG conjugated with horseradish peroxidase (HRP), the secondary antibody from Sigma Aldrich; TMB substrate from Fisher Scientific, New Hampshire, USA were procured.
This study does not involve human subjects or animals and is exempt from ethical approval.
For the development of the ELISA to detect HspX, an in-house procedure was optimized by following a previously described method with minor modifications.11 During the optimization, non-specific binding was ruled out by eliminating various components individually. Experiments were also conducted to optimize the concentration of the monoclonal primary detection antibody (anti-Acr1) and the HRP-conjugated detection antibody. The incubation time and temperature were also optimized.
Cultivation of Mycobacterium bovis BCG Glaxo strain
Mycobacterium bovis was grown in Middlebrook 7H9 broth, prepared by adding 1.25 g of glycerol and 2.35 g of 7H9 powder to 450 mL of distilled water. After filter sterilization, 25 mL of the Middlebrook 7H9 broth base was supplemented with 10% oleic acid-albumin-dextrose-catalase (OADC) and 0.05% Tween 80. The rest of the procedure for developing a growth medium were described previously.12 For determining the exponential growth, the bacteria were allowed to grow, and optical density (OD) was measured spectrophotometrically.
The total bacterial protein from bacteria growing under different conditions was determined by Bradford assay, as previously described.13
Comparing Mycobacterium bovis growth in hypoxic and non-hypoxic environments
After incubation for 7 days, 25 mL of the culture was split equally into two 30 mL tubes (12.5 mL in each). The first tube was incubated in a static incubator in a standing position (hypoxic cells) at 37°C for 4 hours. The other tube was incubated in a shaking incubator (130 rpm) at 37°C for 4 hours. In the settled (standing) tube, the oxygen level had reduced, potentially causing the DosR regulon to mimic the in vivo environment. Bacterial culturing and all tasks were performed aseptically using a Bunsen burner, and the working bench area was decontaminated using 70% ethanol.
Detection of HspX protein in cell culture exposed to nitric oxide
The total protein concentration of the M. bovis BCG Glaxo lysate was determined following exposure to spermine-nitric oxide (Sp.NO) and spermine hydrochloride (Sp.HCl) at a series of exposure times using Bradford assay and the total protein concentration was normalized. It is essential to mention here that Sp.NO is considered a potent nitric oxide donor besides its anti-tumor effect.14 The capture antibody concentration used had been optimized against the HspX protein standard (recombinant HspX protein). First, 100 μL of the anti-Acr1 polyclonal antibody (orb334091, Biorbyt, UK) was dispensed onto a NUNC Maxisorp 96-well microplate at a concentration of 2.5 μg/mL diluted in a coating buffer. The plate was incubated at 4°C overnight. The next day, the wells were washed 3 times in wash buffer, and 100 μL of the recombinant HspX protein (antigen standard) was added in triplicate with serial dilution (15.62, 31.25, 62.5, 125, 250, 500, 1000 pg/mL). In addition, 100 μL of the lysate from the Sp.NO and Sp.HCl exposed samples (separate wells, in triplicate) was added at different concentrations (0.77, 0.83, 0.86, and 0.88 mg/mL) diluted in phosphate buffered saline (PBS), followed by incubation for one hour at room temperature (approximately 25°C). After incubation, the wells were washed 3 times in wash buffer, and 100 μL of the monoclonal detection antibody was added at a concentration of 5 μg/mL in blocking buffer, followed by incubation for one hour at room temperature (approximately 25°C). Then, the wells were washed 3 times in wash buffer, and 100 μL of the goat anti-mouse HRP-conjugated secondary antibody (Sigma Aldrich) diluted in blocking buffer at 1:2000 was added. After incubation, the solution was removed by washing the wells 6 times with the wash buffer, and 100 μL of the TMB substrate (Fisher Scientific, USA) was added to each well, followed by incubation for 15 minutes at room temperature (approximately 25°C), the reaction was stopped by adding 100 μL of sulfuric acid to each well, changing the solution to yellow. Finally, the plate was read using a plate reader at an absorbance of 450 nm.
Lipid body analysis of M. bovis
The cell cultures in the standing and shaking tubes were incubated separately (static and shaking incubators) for 4 hours at 37°C. Tubes containing bacterial culture were then prepared for cell lysis and lipid body analysis. After incubation, 8 mL samples were taken from each culture (standing and shaking), transferred into 2 separates labeled 15 mL tubes, and centrifuged at 500 x g for 10 minutes. The supernatant was removed, and the bacterial pellets in the tubes were stored at -20°C in a freezer. Lipid body analyses were performed using Nile red staining, as described previously.15
Effects of nitric oxide on M. bovis
Mycobacterium bovis was cultured on petri plates for 7 days, until the appearance of a bacterial lawn. Bacteria were harvested using a loop and suspended in 45 mL of Middlebrook 7H9 and then split into 8 of 5 mL aliquots into 30 mL tubes. To induce hypoxia, 4 tubes were exposed to nitric oxide (the donor was Sp.NO), and 4 tubes were exposed to Sp.HCl (control). A 10 mM stock was prepared for Sp.NO and Sp.HCl and diluted to 100-fold to a final concentration of 100 μM. Bacteria were exposed to Sp.NO or Sp.HCl for 0, 1, 2, or 4 hours. For the time course, Sp.NO was added immediately to the cell suspension. All the 30 mL tubes were then sealed and incubated in a shaking incubator (130 rpm) at 37°C for the required exposure time. After completion of the exposure time for Sp.NO and Sp.HCl, 3 mL was taken from each tube (2 mL for cell lysis and 1 mL for lipid body analysis) into a Lysing Matrix (Q-Biogene) tube, centrifuged for 1 minute at 250 x g, and washed 3 times with PBS.
Statistical analysis
Data presented are the average of 3 replicates.
Results
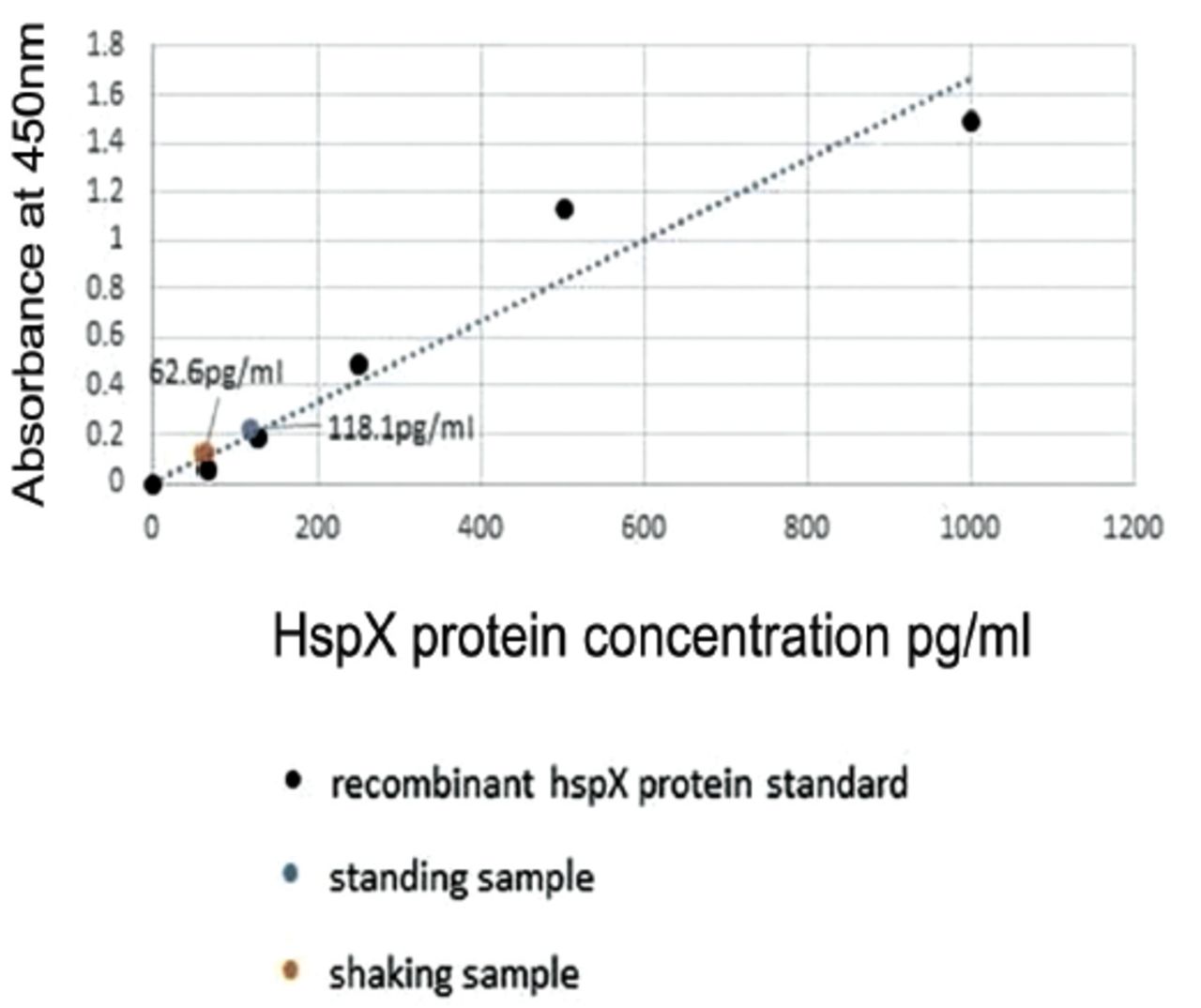
The optimization for the concentration of capture, detection antibodies, and anti-alpha crystallin and recombinant HspX protein samples allowed us to develop a highly reliable in-house detection of this protein. A concentration of 2.5 μg/mL of rabbit polyclonal antibody (capture antibody) was the minimum concentration that yielded positive results and was used as the working concentration in 100 μL volume for the optimized ELISA. The mouse monoclonal detection antibody worked over a broad range (0.31-5.0 μg/mL). The goat anti-mouse IgG conjugated with HRP was highly sensitive and worked even at 1:6000. The minimum detection threshold for HspX was 62.5 pg/mL in the optimized ELISA protocol in this study. The time of incubation that showed maximal results was 3 hours.
The M. bovis BCG Glaxo strain used in this study reached the exponential phase of growth in shaking culture after 7 days with an OD of 0.7 (Figure 1).

- Growth curve for Mycobacterium bovis Bacillus Calmette-Gurin Glaxo strain.
After establishing bacterial growth conditions, exponentially growing M. bovis were subjected to nitric oxide treatment using Sp.NO, which releases nitric oxide in the solution. Spermine hydrochloride was used as a control in this experiment. Treatment with the Sp.NO led to an increase in the total protein concentration of the exponentially growing M. bovis (Figure 2).
- Total protein concentration of Mycobacterium bovis on exposure to spermine-nitric oxide and spermine hydrochloride. Sp.NO: spermine-nitric oxide, Sp.HCl: spermine hydrochloride
A Maxisorp 96-well microplate was set up with the optimized concentrations of the antibodies. Recombinant HspX protein was used as the antigen standard with serial dilutions, with standing and shaking samples at a concentration of 0.22 mg/mL.
The total protein increase under nitric oxide treatment correlated with elevated levels of HspX protein in hypoxic samples in a standing culture, compared to a shaking culture, as detected by the optimized ELISA (Figure 3) and in samples exposed to nitric oxide from Sp.NO compared to the control cells exposed to Sp.HCl (Table 1). Moreover, the HspX protein concentration increased with the increase in exposure time (Table 1). These results indicated that the ELISA protocol developed in our laboratory can accurately detect the levels of HspX protein.

- The absorbance of standing and shaking sample concentrations of Mycobacterium bovis for the HspX protein.
- HspX protein concentration upon exposure to nitric oxide.
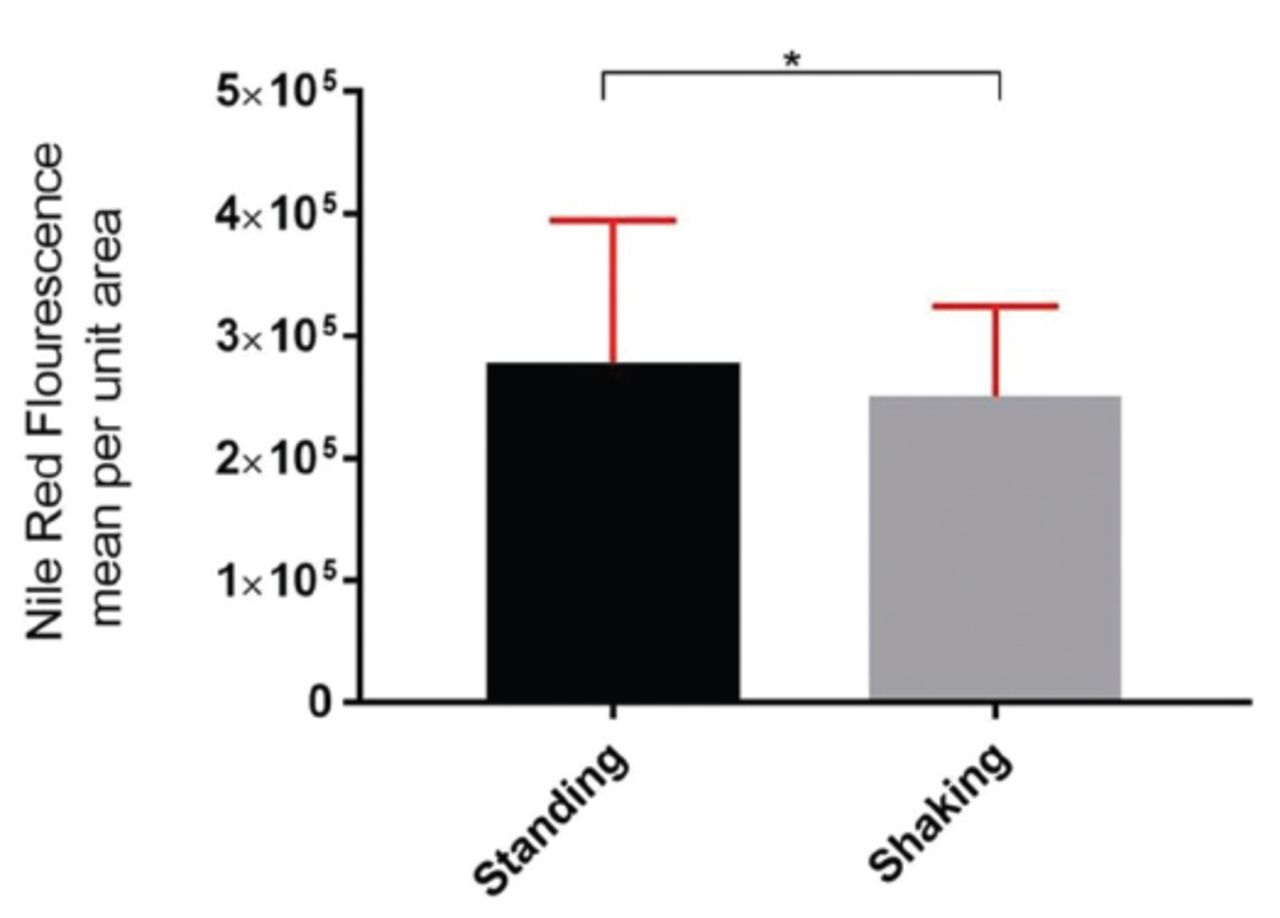
The role of lipid bodies in the pathogenesis of mycobacteria is an active area of investigation. It has been observed that Mycobacterium lipid bodies play an essential role in their survival and contribute to the development of resistance against conventional antibacterial medications.10,16 Staining of bacterial culture from the standing (hypoxic) and shaking culture with Nile red revealed an increase in the level of lipid bodies,15 as shown in Figure 4.
- Fluorescence intensity of Mycobacterium bovis Bacillus Calmette-Gurin Glaxo lipid bodies using 2 different incubation methods.
We also evaluated the effect of nitric oxide on the levels of intracellular lipid bodies in M. bovis culture treated with Sp.NO and Sp.HCl samples and stained with 10 μg/mL of Nile red for 10 minutes, dried and mounted on 10% glycerol in PBS, and sealed with glass coverslips. Fluorescence images were recorded at wavelengths of excitation of 460 nm and emission of 550 nm, using a Nikon Diaphot 300 inverted microscope with a 100-W mercury lamp as a fluorescence source. Figure 5 illustrates that the increase in the lipid bodies in the Sp.NO sample depends on the exposure time.

- Analysis of the fluorescence intensity of Mycobacterium bovis Bacillus Calmette-Gurin Glaxo lipid bodies on exposure to nitric oxide. The production of lipid bodies in the cell culture treated with spermine-nitric oxide (black bar) increased compared with the slight increase in lipid bodies in the cell culture treated with spermine hydrochloride (gray bar).
Discussion
This study aimed to establish a highly sensitive and specific test for the detection of alpha-crystallin (HspX) protein in the in vitro environments. It is crucial to mention here that this protein is also associated with latent mycobacterial infections. The HspX protein in pathogenic mycobacteria provides a survival advantage, and its relative proportion increases during latency and under stressed conditions. It has been reported that almost 25% of the total protein expressed under hypoxic environments in these pathogens is HspX.17 Based on its ubiquitous presence in mycobacteria, it is believed that HspX contributes toward Mycobacterium bacilli survival through inhibiting degradative pathways under stress conditions that can potentially weaken this pathogenic microorganism. Due to the critical role of HspX in Mycobacteria, it has been suggested as the most promising target for developing a vaccine against mycobacterial infections in humans. M. bovis was used in the present study, and functional HspX has been reported in this bacterial species as in Mtb.18
Detection of HspX protein in the sera of TB patients or bacterial culture or intracellularly within the pathogen in vitro has been challenging. In this study, we established an in-house ELISA method for the detection of HspX, and optimized by varying concentrations of each component, (capture and detection antibodies, antigen) and the detection system. This method avoids potential variation between experiments due to the degradation of the components.
Mycobacterium species are among the very few pathogenic bacteria that produce lipids bodies. Limited information is available on the association of the lipids with bacterial survival is concerned. Our findings corroborated previous studies that showed elevated levels of HspX under hypoxic and stress environments. Here, we describe a novel association of elevated HspX levels with lipid bodies. Actively-growing M. bovis culture has relatively lower amounts of lipid bodies than a culture without shaking (mimicking a hypoxic environment). Importantly, treatment of the bacterial solution with nitric oxide was also associated with the increased lipid bodies in M. bovis. This suggests an essential link between HspX and lipid body synthesis in the Mycobacterium species under stressed environments. The pathways involved are yet to be deciphered; however, this finding strongly links HspX elevation with enhanced lipid bodies inside mycobacteria.
In stress environments, higher levels of lipids bodies may be an essential source of energy and carbon. The increased amount of lipids in the cell envelope gives cells the ability to resist the antibiotics, acids, and innate immune responses via the initial replication within the host macrophage.19 This affects the permeability required to transfer the necessary elements for cell growth. As a result, mycobacteria have a slow growth rate and undergo division every 15-20 hours.20 Additionally, owing to the abundance of the lipids in the cellular envelope, cells stick together, forming long cords similar to biofilm formation.21
Moreover, these lipid bodies mainly comprise triacylglycerol (TAG), which is an essential factor during infection. The triacylglycerol synthase 1 (Tgs1) protein plays a vital role in the production of TAG. The Tgs1/Rv3130c gene is one of the DosR regulon members. Triacylglycerol synthase 1 (50.720 kDa) is integrated into the cell wall and cell membrane. Mycobacterium tuberculosis may use Tgs1 to inhibit growth metabolism by altering the acetyl CoA pathway. The exposure to a non-toxic concentration of nitric oxide can reduce the respiration reversibly. Moreover, reduction of oxygen levels or exposure to nitric oxide activates the DosR regulon followed by induction of Tgs1, resulting in an increased production of lipid bodies and antibiotic tolerance.22
Additionally, one of the genes induced by hypoxia in Mtb is the heat shock protein gene (HspX/acr/Rv2031c), which encodes a heat shock protein in the inner membrane. The HspX protein is dependent on the activation of the DosR regulon during the reduction of oxygen. The induction of the DosR regulon follows the production of HspX protein. However, it can be induced independently.23
The findings of this study open new avenues to understand Mycobacterium survival strategies. This will help in the future to devise a strategy for controlling mycobacterial infections among animals and humans.
Mycobacterial infections are highly prevalent in Asia, and HspX is considered the most reliable biomarker. Imported kits always have an issue with the deterioration of temperature-sensitive reagents. Previously, homemade ELISA kits for bacterial infection detection have shown an advantage over commercially imported kits.24 Thus, our study describes the optimization of procedure for detecting HspX protein that has significant implications in understanding the pathogenesis of mycobacterial infections.
Of importance, it is crucial to mention the advantage of the in-house ELISA protocol for the detection of HspX outlined and optimized in this study. Besides significant cost benefits, there is an advantage of accurate detection of this protein linked with mycobacterial pathogenesis. In general, the relative sensitivity and specificity of freshly prepared kits are better than the commercial ones. Sometimes storage conditions may compromise the integrity of temperature-sensitive ingredients. As such, the procedure developed and described in this study offers a unique advantage.
Particular limitations in our study exist that need to be addressed in future studies that include utilizing clinical samples. However, this study provides a fundamental paradigm for further refinements in the procedure established.
In conclusions, our study indicates that in stress environments, such as hypoxia, the elevation of HspX protein is associated with increased lipid body production in M. bovis Bacillus Calmette-Gurin (BCG) Glaxo. These results are consistent with previous studies, which have found that the DosR regulon is induced under hypoxic conditions, resulting in the bacteria entering the dormant state. The dormancy stage is marked by an increase in lipid body production via the simulation of TGS1, which is mainly responsible for producing triacylglycerol. The exposure to NO, as in our study, reduces the oxygen level and induces the DosR regulon, resulting in the production of both lipid bodies and HspX.
Acknowledgment
The author gratefully acknowledges Prince Sultan Military Medical City, Riyadh, Kingdom of Saudi Arabia for the tremendous support. The author would also like to acknowledge Cambridge Proofreading LLC for English language editing.
Footnotes
Disclosure. Authors have no conflict of interests, and the work was not supported or funded by any drug company.
- Received January 11, 2021.
- Accepted May 18, 2021.
- Copyright: © Saudi Medical Journal
This is an Open Access journal and articles published are distributed under the terms of the Creative Commons Attribution-NonCommercial License (CC BY-NC). Readers may copy, distribute, and display the work for non-commercial purposes with the proper citation of the original work.
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.