


Abstract
Objectives: To establish the frequency of Dombrock (DO) blood group genotypes in Western Saudi Arabians and to compare the findings with other populations in the 1000 genomes database.
Methods: This cross-sectional study was carried out between December 2018 and February 2019. A total of 440 blood samples in ethylenediaminetetraacetic acid tubes were collected from unrelated Saudi Arabian blood donors from Jeddah, Saudi Arabia. Deoxyribonucleic acid was extracted, followed by an allele-specific polymerase chain reaction for DO*01 and DO*02 alleles (c.793A>G, rs11276). The allele and genotype frequencies were counted and compared to those in other populations using the Chi-squared test with Bonferroni adjustments.
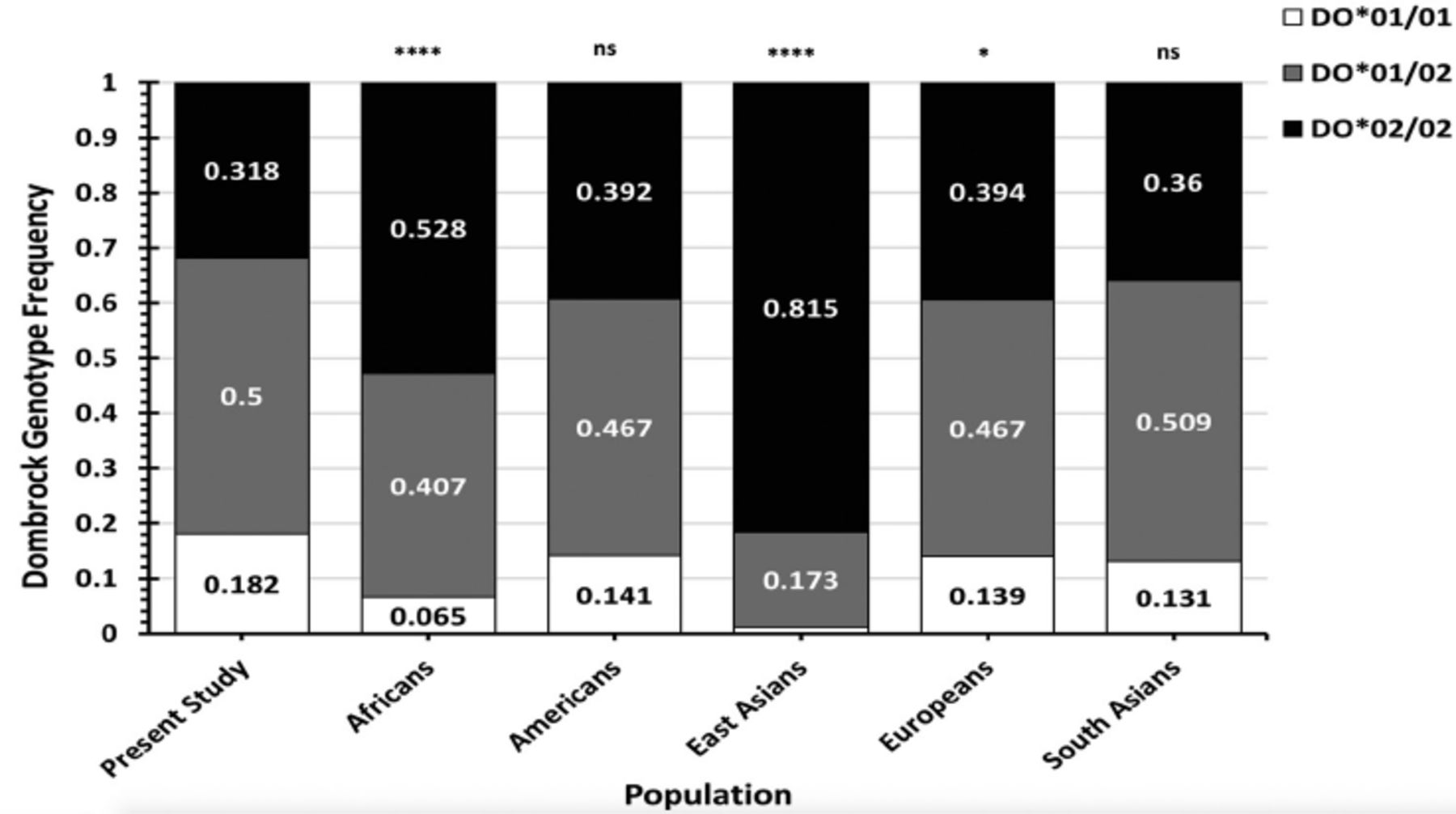
Results: The DO allele frequencies for blood donors from western Saudi Arabia were 0.432 for DO*01 and 0.568 for DO*02. The DO genotype frequencies were 0.182 for DO*01/01, 0.318 for DO*02/02, and 0.5 for DO*01/02. The DO genotype frequencies were similar to Europeans, Americans, and South Asians but significantly different from the genotype frequencies of Africans and East Asians reported in the 1000 genomes database.
Conclusion: Dombrock genotype frequencies in the Western Saudi Arabian population were different from Africans and East Asians but not from Europeans, Americans, and South Asians. This study contributes to a genotyped blood donor database and may advance transfusion safety for patients in western Saudi Arabia.
In 1965, the Dombrock (DO) blood system (International Society of Blood Transfusion [ISBT] 014) was discovered when an alloantibody to the Doa antigen was identified in a female patient.1 Several years later, an anti-Dob antigen was discovered.1 The DO blood group system currently contains 8 antigens harbored by a glycosylphosphatidylinositol (GPI)-linked glycoprotein at the red cell membrane. The Do GPI-linked glycoprotein is encoded by the ADP-ribosyltransferase 4 gene (ART4; DO; CD297) on chromosome 12p13.2-p12.2 The 2 major alleles DO*01 (DO*A) and DO*02 (DO*B) vary by 3 single nucleotide polymorphisms (SNPs), namely, 378>T, 624T>C, and 793A>G in exon 2 of ART4.3 The first 2 SNPs are silent, whereas in the third, the substitution of adenine to guanine at nucleotide 793 causes a single amino acid substitution from asparagine to aspartic acid. Hence, an antithetical relationship between the DO*01 and DO*02 alleles is produced.3
The distribution of DO genotypes varies between populations.4,5 For instance, the homozygous DO*02/02 (DO*B/B) genotype is predominant in East Asian populations at an approximate 80% frequency.4 In Caucasians, the heterozygous DO*01/02 (DO*A/B) genotype is more frequent (46.7%) than the DO*02/02 genotype (39.4%).4 The DO*01/01 genotype is less frequent than the other genotypes, varying between approximately 0.01-0.2 in global populations.4 Interestingly, DO blood group genotyping has been used to differentiate between ethnic groups in Africa, Afghanistan, and Brazil, thereby emphasizing the importance of this blood group as an ethnic marker.6-10
One of the primary functions of a transfusion service is to supply safe erythrocyte units for patients. However, this objective can be challenging, especially for transfusion-dependent patients who may have multiple alloantibodies in their plasma.11 Alloantibodies to DO antigens are involved in hemolytic transfusion reactions and positive direct antiglobulin testing (DAT+) in neonates.1,12 However, the involvement of anti-Do alloantibodies with hemolytic disease of the fetus and newborn has not been reported. Conversely, transfusion reactions caused by anti-Do alloantibodies may be uneventful and are thus under-reported.1 One reason for this issue is the scarcity of reliable, effective monospecific antibodies for classical serological typing. Other reasons include difficulties in alloantibody elution post-transfusion, masking by other alloantibodies, poor Do antigen expression, and low antibody titers.11 Thus, DO allele genotyping has been developed as an alternative strategy to determine patient and blood donor genotypes to improve transfusion safety and provide compatible blood supplies.13,14
Saudi Arabia contains multi-ethnic populations due to its geographical location between Asia and Africa and immigration stemming from religious or economic reasons.15 Importantly, sickle cell disease and thalassemia are highly prevalent in Saudi Arabia, generating crucial challenges in blood banks in providing compatible blood supplies.16 In addition, most Saudi Arabian population studies are restricted to ABO and RhD blood groups.17-19 Limited attempts have been made to establish phenotypes or genotypes for other blood group systems in Saudi Arabia, especially those not routinely screened.20 In particular, the DO blood group in the Saudi Arabian population remains poorly investigated. This study used allele-specific primers and polymerase chain reaction (PCR) to determine the allele and genotype frequencies for DO*01 and DO*02 in blood donors from Western Saudi Arabia. This method was previously validated to detect DO*01 and DO*02 rapidly in blood donors.5 The findings of this study will be used for comparisons with global populations and to establish a DO blood group database in Saudi Arabia.
Methods
Using the 1000 genomes website (https://www.internationalgenome.org/), priori and posteriori sample size calculations were carried out to generate the desired power for statistical comparisons between blood donors from Western Saudi Arabia and other global populations. Initially, the sample size was calculated if the estimated genotype frequencies had deviated from the null hypothesis by approximately 20%, assuming a significance level of 0.05 and a minimum power of approximately 80%. Subsequently, the estimated size was 241 samples. However, a post-hoc analysis of 241 samples demonstrated a low statistical power for analyses. Thus, the sample size was increased to 440 to achieve the desired statistical power of approximately 80%. The number of samples needed for this study was calculated using the G*Power software (version 3.1.9.6., Germany).
Blood samples in ethylenediaminetetraacetic acid (EDTA) tubes were collected from unrelated blood donors from Jeddah, Saudi Arabia. Jeddah city is in Mecca province, Western region of Saudi Arabia (GPS coordinates: 21°32’35.9988’’N and 39°10’22.0044’’E). Donor recruitment was carried out between December 2018 and February 2019. The study was carried out following the Saudi Ministry of Health regulations and the Declaration of Helsinki. Consent for participation in the study was obtained from all blood donors. The Research Ethics Committee at King Abdulaziz University, Jeddah, Saudi Arabia, approved this study (reference number: 2018-021).
The inclusion criteria were as follows: only unrelated and healthy Saudi Arabian blood donors eligible for blood donation following the local standards for blood transfusion were randomly recruited for this study. Non-Saudi Arabian blood donors or donors aged <20 or >58 were excluded from the study.
Deoxyribonucleic acid (DNA) was extracted from whole blood samples in EDTA using a kit from Norgen Biotek Corp (Ontario, Canada). In brief, 150 μl of whole blood was lysed with equal volumes of digestion buffer and mixed with 12 μl of proteinase K. The mixture was incubated at 55°C for 60 minutes. Approximately 200 μl of buffer SK and 300 μl of ethanol were then added to the mixture and vortexed. The mixture was then transferred into a collection tube containing a spin column. The collection tube was centrifuged for 3 minutes at 5,200 g and then washed twice with 0.5 ml of wash solution A. After the washing step, the spin column was transferred into a sterile tube. The spin column was eluted with 200 μl of elution buffer B and centrifuged for one minute at 14,000 g. The previous step was repeated, but the centrifugation time was extended to 2 minutes. Finally, the average DNA concentration was calculated using duplicate reading obtained from the NanoDrop spectrophotometer (NanoDrop Technologies, Delaware, USA) at 260/280 nm.
Blood donors were screened for the DO alleles DO*01 and DO*02 (c.793A>G, rs11276) using PCR-ASP and human growth hormone as an internal control, as previously described.5 Polymerase chain reaction primers are shown in Table 1. The final reaction volume was 25 μl, which contains 1x GoTaq® Green Master Mix (Promega, Wisconsin, USA), 1.0 μM of primers, <250 ng DNA, and DNAse-RNAse free water. Its conditions were divided into 4 phases, as previously described.5 Its reactions were denatured at 94°C for 5 minutes, then 10 reaction cycles at 94°C for 30 seconds, and at 65°C for one minute, followed by 25 reaction cycles at 94°C for 30 seconds, at 60°C for 30 seconds, and at 72°C for 2 seconds. The final extension was carried out at 72°C for 10 minutes.5 Amplified PCRs were then electrophoresed on 2% agarose gels (Sigma-Aldrich, Montana, USA) and visualized using a Bio-Rad documentation system (BioRad, California, USA). Amplicons for both primers were documented as heterozygous DO*01/02, whereas single amplification of DO primer was documented as homozygous for either DO*01 or DO*02. Polymerase chain reactions accompanied by a failed internal control were not documented and repeated. Similarly, PCR amplification with a contaminated negative control was not documented and repeated.
- The sequences of the allele-specific and internal control primers used for genotyping the Dombrock single nucleotide polymorphism c.793A>G in blood donors from the Western region of Saudi Arabia.
Statistical analysis
Dombrock genotype frequencies from PCR were documented using Excel spreadsheets (Microsoft Excel for Mac, version 16.48). Divergences from the Hardy-Weinberg equilibrium were analyzed using the Chi-squared test (χ2). Previous studies on DO genotype frequencies in Saudi Arabian populations were not available. Therefore, the DO genotype frequencies of global populations listed in the 1000 genomes database were used to examine potential similarities with DO genotype frequency in Western Saudi Arabia. For this purpose, χ2 tests for 3×2 contingency tables were carried out. If the p-values obtained from genotype analyses were <0.05, a post-hoc test with Bonferroni correction was carried out to investigate the cause for significance. This test was carried out by partitioning the data as DO*01/01 versus non-DO*01/01, DO*02/02 versus non-DO*02/02, and DO*01/02 versus non-DO*01/02, followed by a χ2 test for each partitioning. Type I errors were corrected by dividing the statistical significance of 0.05 by the number of desired post-hoc analyses (adjusted p-value of 0.0167). Chi-squared and post-hoc analyses were carried out using GraphPad Prism (version 9.1.2., California, USA).
Results
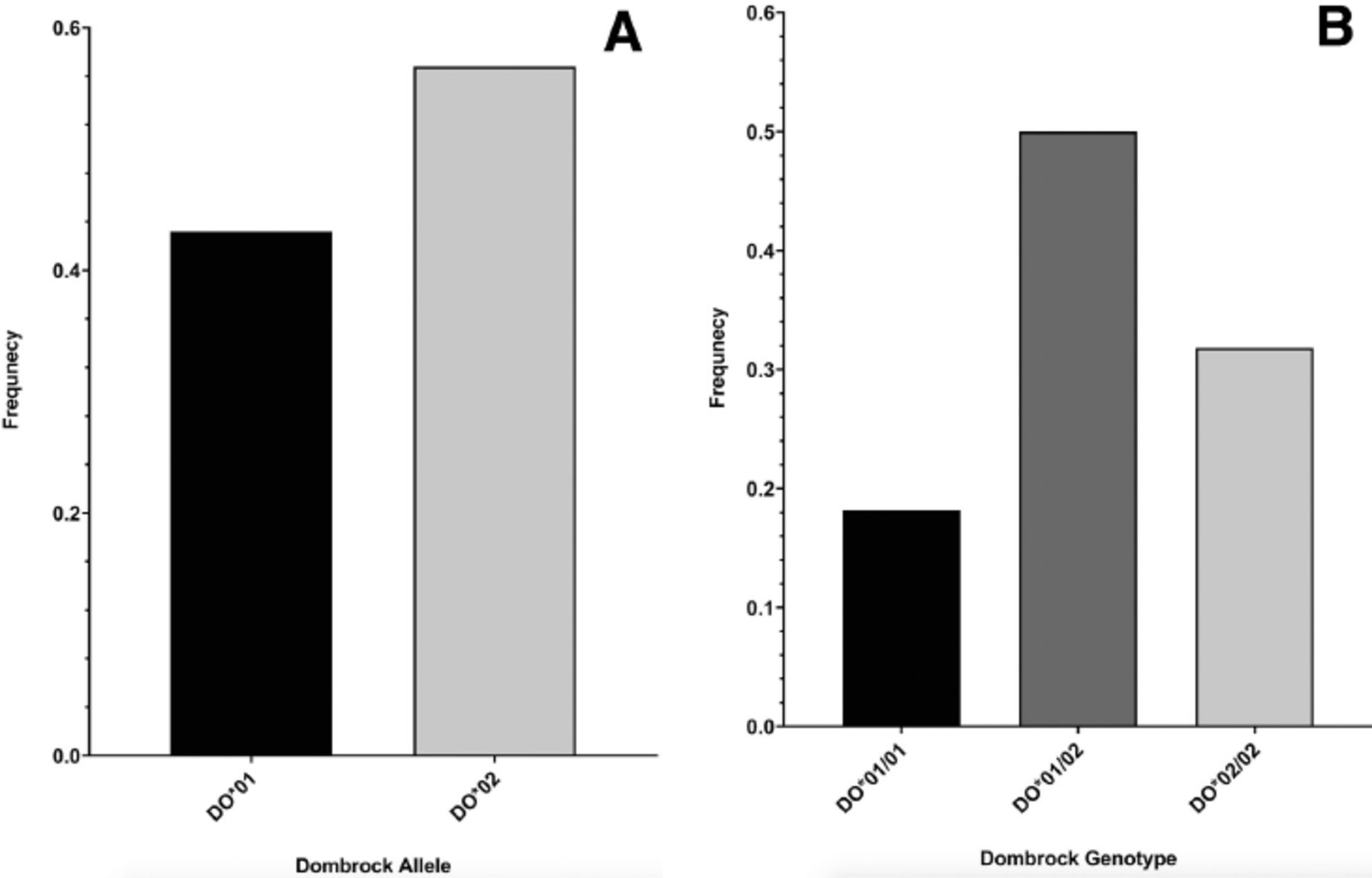
The identified DO genotype and allele frequencies are shown in Figure 1. The DO allele frequency in the blood samples was 0.432 for DO*01 and 0.568 for DO*02. The DO genotype frequency in blood donors was 0.182 for DO*01/01, 0.318 for DO*02/02, and 0.5 for DO*01/02. Genotyping was previously validated.5 The distribution of the DO genotype frequencies was in concordance with the Hardy-Wienberg equilibrium (χ2 [2, N=440]=0.16, p=0.69).

- Dombrock (DO) genotyping results for the c.793A>G (rs11276) single nucleotide polymorphism in blood donors from the Western region of Saudi Arabia. A) Majority of the blood donors have DO*02 (DO*B) allele at approximately 56.8%, whereas the remaining blood donors have DO*01 (DO*A) allele at approximately 43.2%, B) genotyping for the c.793A>G also demonstrated that half of the blood donors from Western Saudi Arabia were heterozygous for the DO blood group (DO*01/02), whereas the homozygous genotype DO*02/02 was present at a frequency of 31.8% and DO*01/01 was present at a frequency of 18.2%
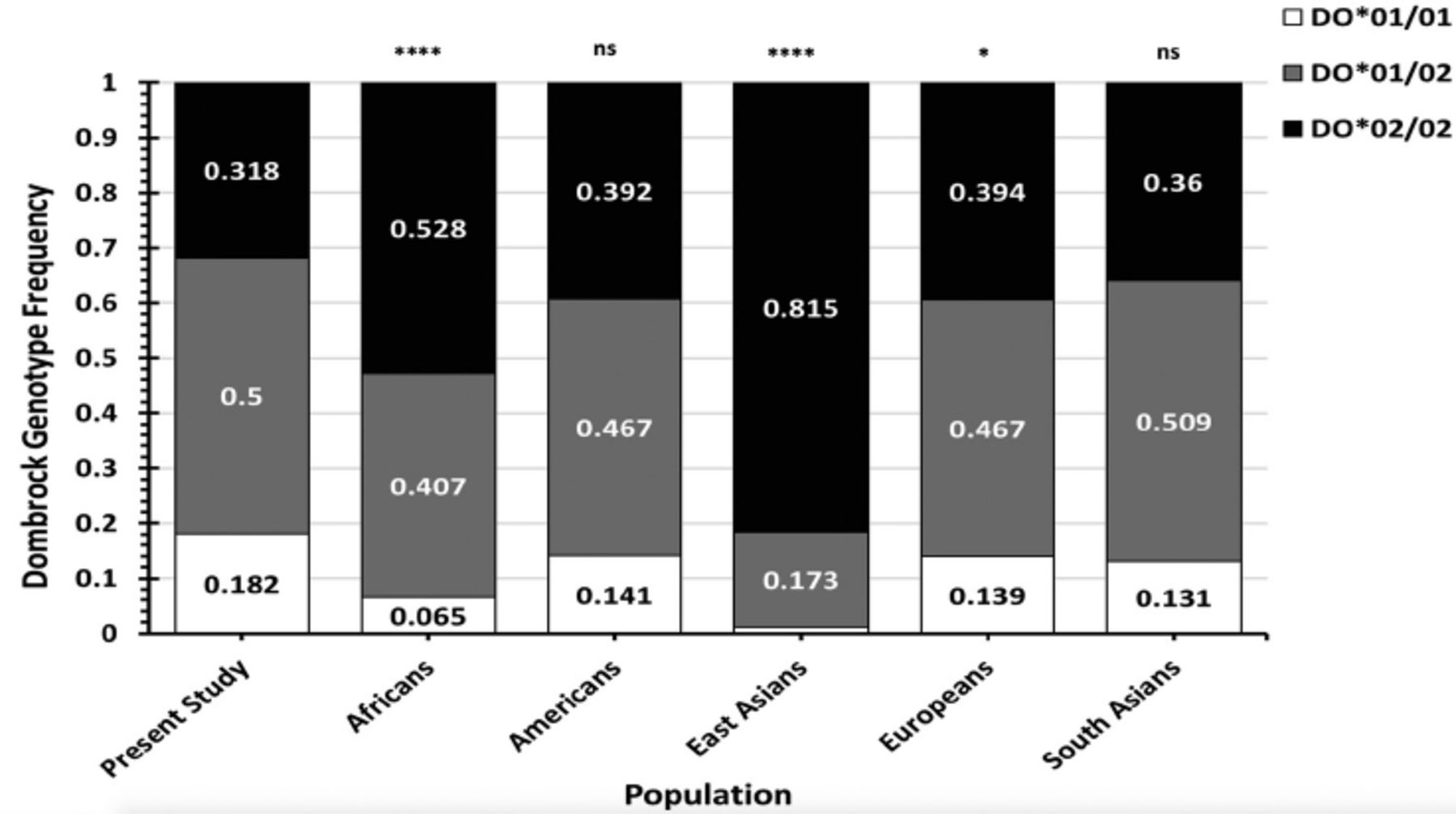
- Stacked bar charts demonstrating the Dombrock (DO) genotype frequencies found in blood donors from the Western region of Saudi Arabia compared with the total DO genotype frequencies of worldwide populations listed in the 1000 genomes database.
Dombrock genotype frequencies were compared to those in global populations from the 1000 genomes database (Figure 2, Tables 2&3). The DO genotype frequencies in Western Saudi Arabians were significantly dissimilar from those in African populations (χ2=63.57, p<0.001) and East Asian populations (χ2=251.4, p<0.001). Conversely, the DO genotype frequencies in Western Saudi Arabians were statistically similar to overall genotype frequencies in American and South Asian populations (p>0.05), except Peruvians from Lima, Peru, (χ2=8.70, p=0.013) and Gujarati Indians from Houston, Texas, United States, (χ2=8.25, p=0.016). Furthermore, the genotype frequencies in Western Saudi Arabians were slightly different from the overall genotype frequencies in Europeans (χ2=6.94, p=0.031). However, further DO genotype frequency analyses in European populations were not statistically different from those in Western Saudi Arabians, except for the Finnish population (χ2=17.97, p<0.01). Post-hoc analyses indicated that genotype DO*01/01 or DO*02/02 frequencies in Western Saudi Arabians were significantly different from most African populations, Peruvians, and Gujarati Indians from United States (p<0.0167). By contrast, all DO genotype frequencies (DO*01/01, DO*01/02, and DO*02/02) in East Asian populations were significantly different from frequencies in Western Saudi Arabians (p<0.01; Table 2).
- The genotype frequencies of the Dombrock blood group polymorphism c.793A>G (rs11276) in Western Saudi Arabians compared with African and East Asian populations listed in the 1000 Genomes database.
- The genotype frequencies of the Dombrock blood group polymorphism c.793A>G (rs11276) in Western Saudi Arabians compared with those in American, European, and South Asian populations listed in the 1000 Genomes database.
Discussion
Analyzing blood group genotype frequencies can be used as markers for the identification of variations between populations and local ethnic admixtures and also for the improvement of the safety of transfusion practices.6,14 This study aimed to analyze DO allele and genotype frequencies in Western Saudi Arabia to establish a blood donor database. The study’s data confirmed that the most common genotype in 440 Western Saudi Arabian blood donors was the heterozygous DO*01/02 (50%) genotype, followed by the DO*02/02 (31.8%), and DO*01/01 (18.2%) genotypes. These frequencies were similar to the majority of European, American, and South Asian populations (p>0.05) but significantly different from East Asians and most African populations.4 This significance was due to differences in DO*01/01 and DO*02/02 genotype frequencies. The present study analyses also indicated that DO genotype frequencies in Western Saudi Arabians, African Americans in the South-West United States, and Mende populations in Sierra Leone were statistically insignificant. Nonetheless, the DO*02/02 frequency in Western Saudi Arabians was nearly significantly different from these African sub-populations. In addition, the number of blood donors examined in these sub-populations was <100, potentially increasing the risk of statistical error. Thus, increasing blood donor numbers for DO genotype testing in these populations may provide statistical differences to Western Saudi Arabian data. Overall, the genotype frequency of most African populations was significantly different from Western Saudi Arabians, despite the close geographical proximity. Dombrock genotype frequency differences were more apparent between Western Saudi Arabians and East Asians, where DO*02/02 was more frequent in East Asians (approximately ≥80%) compared with that in other populations, including Western Saudi Arabians.4 In addition, the overall DO genotype frequencies in Europeans were significantly different from Western Saudi Arabians. However, when the DO genotype frequencies reported in the Finnish population were excluded from analysis, the DO genotype frequencies were statistically similar between Western Saudi Arabians and the remaining European populations (p>0.05). Collectively, the blood group heterogeneity frequencies in Western Saudi Arabians may be due to immigration, as previously suggested.15 A recent study on the distribution of major blood groups in Saudi Arabians reported that the frequency of some blood group phenotypes was similar to that in Africans or Caucasians, whereas other phenotype frequencies were distinctively high in Western Saudi Arabians.21 Another study demonstrated that the molecular background of the Fynull phenotype in Western Saudi Arabians was similar but at a lower frequency than that reported in Africans.20 Hence, Western Saudi Arabians may contain a distinct genetic pool separate from all populations. Further genotyping of other blood groups in the Saudi Arabian population may provide additional insights into immigration and its influences.
Alloantibodies to DO blood groups are implicated in transfusion reactions and positive DAT in neonates, thereby underscoring its clinical significance.1,12 In blood banks, routine screening and identification of alloantibodies to blood group antigens are regularly carried out. Nonetheless, the identification of blood group antigens or alloantibodies in patients’ or donors’ blood using standard serological techniques may be challenging, as reliable anti-Do anti-sera for such identification is lacking.1 Importantly, anti-Do alloantibodies may be masked with the existence of alloantibodies to other blood group antigens, adding further complexity to transfusion practices.1 Hence, alloimmunization or transfusion reactions due to Do mismatch may be under-reported in Western Saudi Arabia.11 In Saudi Arabia, the rate of patients with sickle cell disease and thalassemia requiring multiple transfusions is high.16 This issue is particularly important as these patients require multiple transfusions, increasing the risk of alloimmunization and transfusion reactions. In a recent study, alloantibodies to major blood groups were identified in Saudi Arabia in patients with sickle cell disease and thalassemia.22 Therefore, the authors of the study have recommended routinely extended phenotyping to reduce the transfusion of mismatched units and prevent alloimmunization and potential transfusion reactions.22 For such risk groups, the genotyping of clinically significant blood groups may increase blood transfusion safety metrics and improve the reliability of anti-sera used by blood banks, especially for anti-sera with known difficulties, such as anti-Do antibodies.14,23 In addition, extended genotyping data for patients with sickle cell disease and thalassemia in Saudi Arabia remain to be elucidated. However, mismatch and alloimmunization to other major blood groups may exist. A Brazilian study using DNA array analysis and electronic matching for compatibility indicated that less than one-tenth of blood donors (approximately 6.3%) were compatible with patients with sickle cell with both Do (a-) and other antigen-negative phenotypes.14 Therefore, future studies focusing on genotyping for patients with sickle cell disease and thalassemia and compatible donors may provide insights on their alloimmunization profiles and implement strategies to increase blood transfusion safety.
Study limitations
Other SNPs of the DO blood group system were not analyzed. However, these SNPs are rare, as previously reported.24,25 In addition, this study was limited to Western Saudi Arabia. Hence, the reported data cannot be generalized to all Saudi Arabians. A nationwide genotyping study is required to analyze the frequency of different blood groups in the Saudi Arabian population to increase our understanding. Such studies may highlight regional differences across the country, establish a robust database for clinical applications, and ultimately improve patients’ safety.
In conclusion, DO genotype frequencies in the Western Saudi Arabian population were different compared with those in Africans and East Asians but not with those in Europeans, Americans, and South Asians. Importantly, this study has contributed data to a genotype blood donor database that can be used to improve transfusion safety for patients.
Acknowledgment
The author gratefully acknowledge Scribendi (https://www.scribendi.com) for English language editing.
Footnotes
Disclosure. Author has no conflict of interests, and the work was not supported or funded by any drug company.
- Received November 9, 2021.
- Accepted January 17, 2022.
- Copyright: © Saudi Medical Journal
This is an Open Access journal and articles published are distributed under the terms of the Creative Commons Attribution-NonCommercial License (CC BY-NC). Readers may copy, distribute, and display the work for non-commercial purposes with the proper citation of the original work.
References
In this issue




{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.