


Abstract
Myocardial ischemia and reperfusion (I/R) injury is a pathological condition characterized by an initial restriction of blood supply to the heart followed by the subsequent restoration of perfusion and concomitant re-oxygenation. This condition may cause heart injury and contribute to morbidity and mortality. Although tremendous advances have been made in understanding the mechanisms of myocardial I/R injury, the translation of these findings into the clinical setting has been largely disappointing. MicroRNAs (miRNAs) are endogenous, single-stranded, non-coding RNAs ranging from 18-24 nucleotides in length. They are highly conserved and ubiquitously expressed in all species, which control diverse cellular functions by either promoting degradation or inhibiting target mRNA translation. In particular, a multitude of studies demonstrated miRNAs played an important role in acute and chronic cardiovascular disease processes. In this review, we focus on miRNAs and summarize the latest insights on the role of the specific miRNAs in myocardial I/R injury.
Heart damage caused by ischemia reperfusion (I/R) injury represents a serious event, which often leads to deterioration or even loss of heart function, limits the benefits of reperfusion after acute myocardial infarction (AMI) and has caused a serious global health problem.1 Myocardial I/R injury is a complex pathophysiological event, thanks to the restoration of blood flow to the occlusion vessel.2 Serious acute and chronic heart damage is aggravated because of reactive oxygen and nitrogen species and inflammatory reactions.3 Indeed, a wide range of pathological processes contribute to myocardial I/R injury (Figure 1).3 The hypoxia and the following oxidative stress result in protein modifications, lipid oxidations, and DNA breakage, triggering a chain of deleterious responses that affect all major extra-and intra cellular tissue components: endothelial dysfunction, neutrophil adherence to endothelium, and trans-endothelial migration, the release of inflammatory mediators, cellular calcium overload, and eventually cell death. These events are the underlying mechanism of acute I/R damage and dysfunction in the heart. However, although tremendous advances have been made in understanding the mechanisms of myocardial I/R injury, the translation of these findings into the clinical setting has been largely disappointing.3 Despite the recent advances, the underlying molecular signaling between cellular components, extracellular matrix, and tissue vascularization associated with myocardial I/R injury are far from being completely understood. MicroRNAs have been implicated as transcriptional regulators in a wide range of biological processes determining cell fate, stress response, proliferation, or death.4-6 Recently, several studies have suggested that miRNAs contribute to I/R injury by altering key signaling elements; thus, making them potential therapeutic targets. Our data also has demonstrated that miRNA-22 could inhibit apoptosis of cardio myocytes through one of its targets cAmp response element binding (CREB) binding protein (CBP), which may constitute a new therapeutic target for the prevention of myocardial I/R injury.7 Here, we highlight the latest advances in the identification of myocardial ischemia-associated miRNAs and their functional significance in the modulation of I/R-induced cardio myocyte death (necrosis/apoptosis), myocardial inflammation, fibrosis, compromised contractile function, and neoangiogenesis (Table 1).
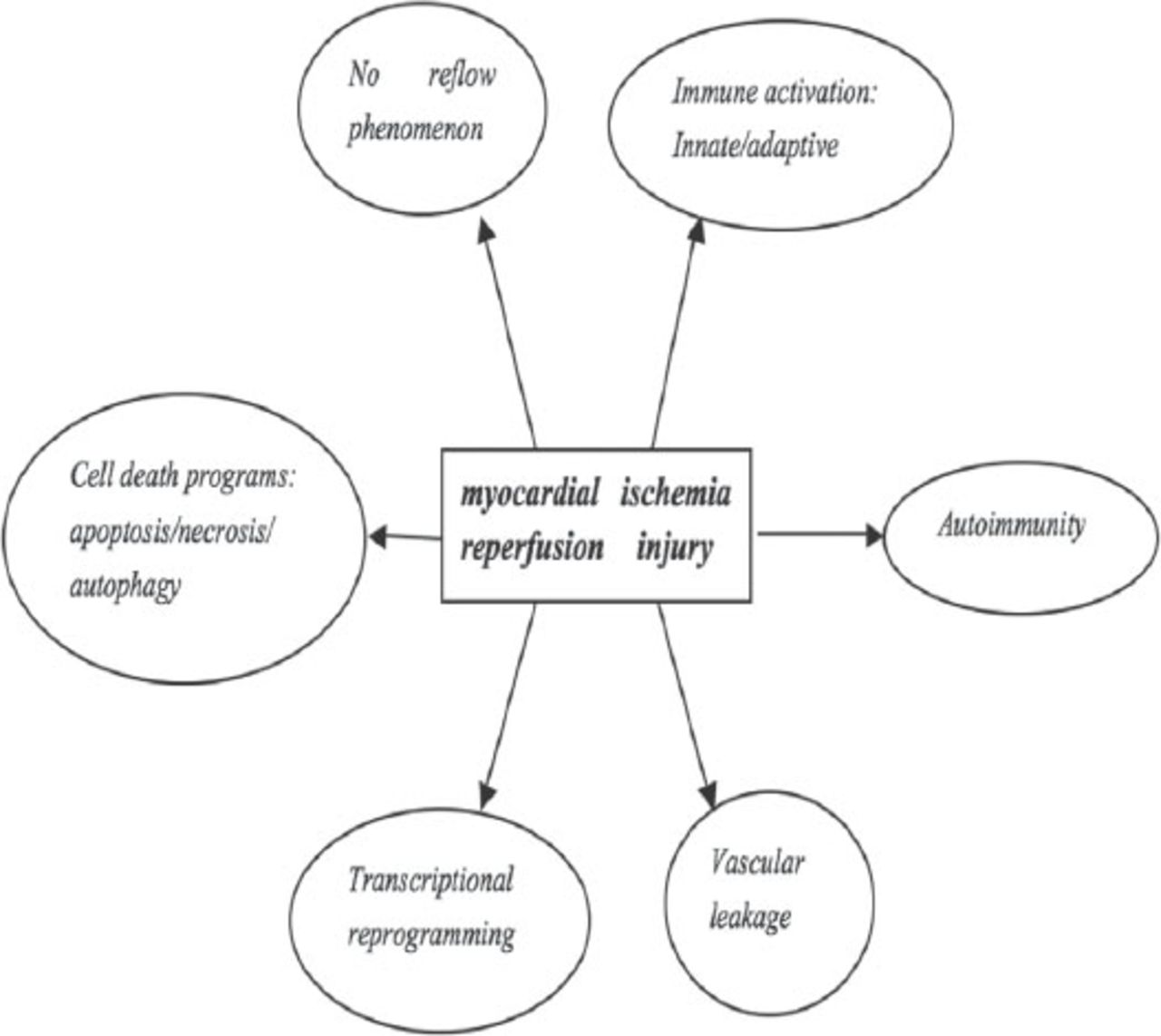
Pathological processes contribute to ischemia and reperfusion associated tissue injury. Copyright permission from Eltzschig HK, Eckle T. Ischemia and reperfusion-from mechanism to translation. Nature Medicine 2011; 17: 1391-1401.
The role of microRNAs in myocardial ischemia reperfusion injury.15
Source and function of microRNAs
MicroRNAs are a class of endogenous, small (~22 nucleotides [nt] in size), noncoding single-stranded RNAs.4 The genes are located in the intron, non-coding exon, and intergenic regions of genomes, and initially transcribed by polymerase II into primary miRNAs (pri-miRNAs).7 A stem-loop pre-cursor miRNA (pre-miRNA, ~70 nt in size) would be formed after the pri-miRNAs subsequently are cleaved by a microprocessor complex composed of Drosha and Pasha. Later, the formed pre-miRNAs would be transported into the cytoplasm. After this process, Dicer would cleave them into mature miRNAs.8 The mature miRNAs can be incorporated into the RNA-induced silencing complex (RISC) and negatively regulate gene expression through binding on the untranslated region (UTR) of target mRNAs.9 The miRNAs could regulate genes involved in a series of diverse biological processes, including development, differentiation, inflammation, stress response, angiogenesis, adhesion, proliferation, and apoptosis by this mechanism.10,11
Expression of microRNAs in myocardial ischemia reperfusion injury
He et al12 detected the miRNAs expression of the myocardial ischemia Sprague-Dawley (SD) rats after reperfusion in one hour, and 3 hours (I/R, 1h/3h) and found that 16 kinds of miRNAs were significantly disordered, of which 10 were up-regulated, and 6 were down-regulated. When reperfusion in 30 minutes or in 24 hours (I/R, 30min/24h), only the miRNA-1, miRNA-126, miRNA-208 were up-regulated, but miRNA-21, miRNA-133, and miRNA-195 reduced.13 At the same time, the myocardial ischemia FVB/N mice (offers a system suitable for transgenic experiments and subsequent genetic analyses) after reperfusion in 30 minutes or in 24 hours (I/R, 30min/24h) miRNA-320 was also observed to down-regulated, but miRNA-7, miRNA-21, miRNA-146b, and miRNA-491 increased.14 Thus, we could draw a conclusion that the miRNA are expressed in myocardial I/R depending on the dynamic nature and condition. The miRNAs disorder expression in early reperfusion were related to myocardial cell death and oxidative stress. Therefore, analysis of the expression and function of myocardial I/R related miRNAs may be better to illustrate the mechanism of myocardial I/R injury.15
Key microRNAs, which play an important role in myocardial I/R injury. 1) Protective microRNAs in myocardial I/R: miRNA-126, miRNA-133, miRNA-144, miRNA-145, miRNA-199, miRNA-210, miRNA-214, miRNA-494, miRNA-451, and miRNA-499
The miRNA-126, as an angiogenic factor, is highly expressed in the heart, which could be maintained to promote blood vessel growth by enhancing the proangiogenic actions of vascular endothelial growth factor (VEGF) and fibroblast growth factor (FGF), and promoting blood vessel formation by repressing the inhibitor Spred-1 and vascular cell adhesion molecule 1 (VCAM-1).16-18 The expression of miRNA-126 increased after myocardial I/R. When miRNA-126 was knockout, the survival rate is significantly reduced.19 At the same time, a research by Zernecke also has proven that miRNA-126 can also inhibit apoptosis by activating the CXC-Motiv-Chemokin12 (CXCL12) chemokine receptor chemokine receptor 4 (CXCR4).19 Thus, currently we have evidence that miRNA-126 is a protective miRNAs in myocardial I/R enhancing angiogenesis and inhibiting apoptosis.
The miRNA-133 is expressed in cardio myocytes. miRNA-133 levels are reduced after myocardial I/R injury.20 When researchers up-regulated the levels of miRNA-133, cell apoptosis was reduced notably and the viability of cardiomyocytes (H9C2) cells after exposure to hydrogen peroxide (H2O2) was also increased. However, after the expression of miRNA-133 was down-regulated by oligonucleotide, the results were opposite. The mechanism may be related to its targets; namely, miRNA-133 would cut the pro-apoptotic gene caspase-9 expression.21 Previous studies21 have shown that increasing miRNA-133 levels would reduce myocardial I/R injury.
The miRNA-144 and miRNA-451 were shown to protect cardio myocytes from apoptotic cell death in vitro.22,23 Conversely, the knockdown of endogenous miRNA-144 and miRNA-451 augmented cell death. The CUG triple repeat-binding protein2, which has been described to regulate the translation of COX-2, was identified as a bona fide target of these 2 miRNAs. The effects of miRNA-144 and miRNA-451 modulation were blocked by simultaneous inhibition of COX-2 by specific inhibitors, NS-398 or DUP-697, further high lighting the importance of this regulatory pathway in the protection from myocardial I/R injury.22,23
An increasing number of studies have declared that up-regulating the intracellular divalent calcium ions (Ca2+) concentrations are closely associated with the activation of calcium/calmodulin-dependent protein kinase (CaMKII), which up-regulates the expression of apoptotic genes. Recently, Cha et al24 revealed that miRNA-145 inhibited Ca2+ overload and Ca2+ -related signals by targeting CaMKIIdd, and the over-expression of miRNA-145 protects against reactive oxygen species (ROS)-induced cardio myocyte apoptosis. The miRNA-199 family includes miRNA-199a-1, miRNA-199a-2, and miRNA-199b. Rane et al25 reported that under anoxic conditions, the cells may rapidly reduced cardiac miRNA-199a expression by a post-transcriptional mechanism. Down-regulation miRNA-199a could induce apoptosis during hypoxia by increasing the expression of hypoxia inducible factor-1α and sirtuin 1 (srt1). Some researchers24,25 also supported that knockdown miR-199a before hypoxia may play a role in pre-treatment and protection.
Just like miRNA-126, miRNA-210 has been shown to possess pro-angiogenic properties.26 Up-regulation of miRNA-210 would increase the migration of vascular endothelial cells. On the contrary, when in hypoxia circumstances, decreasing the expressions of miRNA-210 would inhibit endothelial cell migration and cell survival. miRNA-210 could promote hypoxia miRNA-210-mediated cell survival, migration and angiogenesis by inhibiting ephrin-A3 gene.26 Mutharasan et al27 also found that miRNA-210 would be over-expressed in myocardial hypoxia factor-dependent and independent pathways, which would inhibit oxidative stress-induced apoptosis by influencing the formation of ROS. In addition, Kim et al28 revealed that miRNA-210 had a function to promote cells survival by blocking caspase-8 related protein-2 after myocardial I/R. In summary, the above related studies have shown that miRNA-210 myocardial protection could be used as a new way to treat ischemic heart disease.
miRNA-214 was shown to be highly increased in the models of cardiac hypertrophy and heart failure.22 Genetic deletion of miRNA-214 aggravated myocardial I/R injury, increased apoptotic cell death, promoted the excessive accumulation of fibrotic tissue, and resulted in a loss of contractility. While, when researchers up-regulated the levels in a variety of cardiac disease models, miRNA-214 would play a protective role. Up to now, the common concept of the mechanism of this biological function is related to its role of inhibiting the expression of the Ca2+-handling molecule Na+-Ca2+ exchangers1 (NCX1), as well as CaMKII, and cyclophilin D.29,30
miRNA-499 was found at intron and highly expressed in the heart.22,31 The expressing levels of myocardial miRNA-499 were identified as being down regulated after cardiac I/R injury. The direct targets of this miRNA were α- and β-isoforms of the calcineurin catalytic subunit. Targeting of these 2 subunits resulted in a reduction of mitochondrial dynamin-related protein-1 expression through blunted calcineurin-mediated dephosphorylation of DRP1 and subsequently an aggravation of cardio myocyte apoptosis. In addition to this, researchers have proven that the transcriptional repression of miRNA-499 during I/R injury was shown to be mediated by enhancing the expression of p53.31 AntagomiR knockdown miRNA-499 could also increase collagen deposition, promote myocyte hypertrophy, injury systolic function, and aggravate I/R cardiac remodeling.22 These results indicate that increasing the levels of cardiac miRNA-499 not only could reduce I/R injury, but also delay the AMI remodeling process.22,31
2) Opposite effects of microRNAs in myocardial I/R: miRNA-1, miRNA-15, miRNA-92a, and miRNA-320
miRNA-1 and miRNA-133 (discussed above) were located in the same chromosomal loci, transcribed to be mature simultaneously and independently, but with different biological functions. Studies have shown that miRNA-1 and miRNA-133 play a different role in stress-induced cardio myocyte survival, miRNA-133 inhibits apoptosis (discussed above), whereas, miRNA-1 promotes apoptosis.21 Increasing the miRNA-1 levels of H9C2 rat embryonic ventricular myocytes or rat myocardial cells could stimulate stress-induced apoptosis by down-regulating the expression of several anti-apoptotic genes such as Hsp60, Hsp70, IGF-1, and Bcl-2.21 In addition, injecting miRNA-1 may exacerbate cardiac arrhythmias. The above results suggest that miRNA-1 could lead to cell apoptosis by affecting anti-apoptotic genes.
The miRNA-17-92a gene cluster (encoding miRNA-17, miRNA-18a, miRNA-19a/b, miRNA-20a, and miRNA-92a) levels were increased 24 hours after coronary artery occlusion in mice. This gene cluster could affect angiogenesis dependent on cellular environment.32 Bonauer et al33 showed that antagomir-92a improved left ventricular function, reduced myocardial infarction size, reduced apoptosis, and increased the number of new blood vessels. Researchers33 revealed that the mechanism of this process was due to the effect of this gene cluster on a wide of target genes, such as Srt, ITGA5, S1P1, MKK4, and eNOS. Thus, inhibiting miRNA-17-92a may potentially reduce infarct size after myocardial infarction. The miRNA-15 was also found to be dysregulated in animal models after I/R injury. When inhibiting the expression of miRNA-15 with anti-miR-15, it would protect cardio myocyte cells from hypoxia-induced death in vivo.21,34 The expression of miRNA-320 decreased in myocardial I/R injury.35 Over-expression of cardiac-specific miRNA-320 transgenic mice increased the area of AMI and apoptosis. Antagomirs miRNA-320 reduced the myocardial infarct size. Transfection of miRNA-320 into cardiac microvascular endothelial cells inhibited angiogenesis by inhibiting IGF-1 expression.21 Therefore, silencing miRNA-320 expression by antagomirs could reduce myocardial cell death, and increase neovascularization.
3) Double-edged microRNAs in myocardial I/R: miRNA-21, miRNA-24, and miRNA-29
In recent years, several miRNAs have been proven to play a dual role in the pathomechanism of myocardial I/R injury. miRNA-21 was just one of these miRNAs.36 miRNA-21 was found to be specifically localized to fibroblasts in the infarct region of the heart by immunohistochemistry techniques. Phosphatase and tensin homolog deleted on chromosome ten (PTEN) identified as a direct target was found to regulate the downstream expression of MMP-2.37 miRNA-21 was indispensable for myocardial protection in that setting, and was found to reduce hypoxia/reoxygenation-induced cell apoptosis via regulation of its target gene programmed cell death protein 4 (PDCD4).37 In addition to this, some scholars believed that miRNA-21 could inhibit some other pro-apoptotic genes involved in pro-survival pathways, such as programmed cell death protein 4 (PCDF4) and fas ligand (FasL).38,39 Over-expression cardiac-specific miRNA-21 transgenic mice could reduce myocardial injury,38 and antagomir silence miRNA-21 could increase H2O2-induced myocardial necrosis and apoptosis.39 Studies have shown that miRNA-21 could also promote the survival of fibroblasts and lead to myocardial fibrosis by regulating protein sprouty homolog 1 (Spry1).40 However, another study41 found that over-expression of miRNA-21 reduced endothelial cell proliferation, migration, and angiogenesis by the regulation of ras homolog family member B (RhoB). Thus, inhibiting the expression of miRNA-21 also could promote coronary angiogenesis and restore blood flow, which plays a role in myocardial protection after myocardial infarction. In short, miRNA-21 played a dual role in this area by influencing endothelial cell proliferation, migration, formation of new blood vessels, and cardiac cells survive through its certain targets.42 Like miRNA-21, miRNA-24 plays a similar role in this process.17 miRNA-24 is enriched in cardiac endothelial cells and is up-regulated after I/R injury. Blocking endothelial miRNA-24 limited the injury via prevention of endothelial apoptosis and enhancement of vascularity,43 which led to preserved cardiac function and survival. However, miRNA-24 expression is down-regulated in the ischemic border after I/R. Delivery of miRNA-24 into the infracted hearts of mice inhibited cardio myocyte apoptosis, attenuated infarct size, and reduced cardiac dysfunction.44 This anti-apoptotic effect on cardio myocytes in vivo is partly mediated by the B-cell lymphoma (Bc)l-2 gene. Thus, miR-24 played a dual role in vascular endothelial cells and cardiac myocytes. It is crucial to intervene miR-24 at the right time. Another dual microRNA is miRNA-29. van Rooij et al45 found that them iRNA-29 family (miRNA-29a, b, c) were down-regulated after I/R injury. miRNA-29 can regulate pro-fibrotic proteins gene expression.46 Antagomir, (a kind of inhibitor, which can prevent miRNA and its target genes mRNA complementary pairing) silence miRNA-29 increasing the expression of collagen. In fibroblasts, over expression of miRNA-29 gene transcription could reduce collagen. In addition, miRNA-29 also negatively regulated anti-apoptotic genes such as Tcl-1, Mcl-1, YY1, p85α, CDC42, and DNMT3.36,46 Thanks to its protective mechanism above, it inhibited I/R injury-induced myocardial cell death, and reduced myocardial remodeling. However, at the same time, it may also affect fibroblasts, which would suppress the treatment after myocardial I/R injury. Therefore, the expression of miRNA-29 in the heart has a double-edged effect.36
In conclusion, myocardial I/R injury is determined by numerous mediators and signaling pathways including myocyte death (necrosis, apoptosis, and autophagy), myocyte hypertrophy, cardiac fibrosis, and impaired angiogenesis. In recent years, a growing number of miRNAs related to myocardial I/R injury were found. Identification of I/R-associated miRNAs is important as miRNAs targeting is becoming a topic of interest that could provide new ideas for the treatment of myocardial I/R injury. The role of different miRNAs are very different in this process. Briefly, miRNAs including miRNA-126, miRNA-133, miRNA-144, miRNA-145, miRNA-199a, miRNA-210, miRNA-214, miRNA-494, miRNA-451, miRNA-499 are protective after myocardial I/R injury. While miRNAs including miRNA-1, miRNA-15, miRNA-92a, and miRNA-320 have the opposite effects. Complexly, some other miRNAs such as miRNA-21, miRNA-24, and miRNA-29 have a dual role. Modulation of miRNA levels directly or indirectly provides a therapeutic advantage to interfere with myocardial I/R injury, and the complexity of miRNA targets might yield different consequences in various cell types within the heart. However, up to now, the mechanism of the various specific miRNAs in myocardial I/R injury remained to be further clarified. Hence, to obtain knowledge of the potential target for the treatment of myocardial I/R injury, further study is required to ascertain the many functions of miRNAs in normal physiology and disease states.
Related Articles
Fan ZX, Yang J. Microribonucleic acids and vascular restenosis. Saudi Med J 2014; 35: 796-801.
Xu XH, Liu Y. Study of the correlationship link between microRNAs and nasopharyngeal carcinoma. Saudi Med J 2014; 35: 329-335.
Zhou Q, Deng Q, Huang J, Chen JL, Hu B, Guo RQ. Evaluation of left ventricular mechanical dyssynchrony in patients with heart failure after myocardial infarction by real-time three-dimensional echocardiography. Saudi Med J 2012; 33: 256-261.
Acknowledgment
We would like to thank Chao-Jun Yang for designing this work.
Footnotes
Disclosure. This work was supported by the National Natural Science Foundation of China (Grant No. 81200088 and 81470387), the Natural Science Foundation of Yichang City, China (Grant No. A12301-01), and Hubei Province’s Outstanding Medical Academic Leader Program, Hubei, China. Authors have no conflict of interests and the work was not supported or funded by any drug company.
- Copyright: © Saudi Medical Journal
This is an open-access article distributed under the terms of the Creative Commons Attribution-Noncommercial-Share Alike 3.0 Unported, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
References
In this issue




{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.