


Abstract
Objectives: To investigate the effects of venlafaxine (VEN) on the relapse of methamphetamine (METH)-induced conditioned place preference (CPP) in rats.
Methods: This study was conducted at the Department of Pharmacology and Toxicology, and the Addiction and Neuroscience Research Unit, College of Pharmacy, Taif University, Taif, Kingdom of Saudi Arabia between May 2017 and October 2018. A total of 32 male Wistar rats were used in this study. Rats were divided to receive either METH or saline during the conditioning phase. Following the acquisition of METH-induced CPP, the animals were randomly divided to receive saline or VEN for 21 days instead of METH during the extinction training. The reinstatement was initiated by re-exposure to a single dose of METH to test any anti-reinstatement effects of VEN.
Results: Two-way repeated measures Analysis of Variance (ANOVA) (time × chamber) was used to analyze time spent in conditioning chambers, while distance traveled was analyzed using one-way ANOVA. Saline or VEN, when administered alone, did not affect CPP or locomotor activity results. A priming intraperitoneal injection of METH reinstated CPP in the animals treated with saline during the extinction phase. Interestingly, VEN treatment blocked METH-induced CPP.
Conclusion: Venlafaxine effects on the reinstatement of METH-induced CPP are not likely due to nonspecific effects on locomotor activity. This beneficial effect of VEN on relapse to METH-induced CPP could be due to its antidepressant effects. Venlafaxine can thus be a potential therapeutic option in the treatment of relapse to METH-seeking behaviors.
Drug addiction is a critical health issue in Saudi Arabia and worldwide. Amphetamine is the most commonly used psychostimulant. According to the world drug report, Saudi Arabia reported the highest amphetamine-type stimulant seizure (11 tons) in 2011,1 and methamphetamine (METH) is one of the most commonly abused stimulants worldwide. This indicates a serious and critical health problem due to the addictive properties of METH. Relapse to drug use is the most critical challenge in treating addiction. Several reasons for relapse to drug use have been documented, and the most important reasons are the exposure to stimuli mimicking the environment of previous drug use, the reuse of the drug, and stress.2,3 Therefore, preventing relapse to drug use is the essential goal in the treatment of any drug use disorder. Thus, the focus of this study is to investigate measures that prevent relapse to METH use. Venlafaxine (VEN), a well-known antidepressant, showed efficacy in the treatment of cocaine dependence in humans.4 More precisely, VEN has been reported to restore neurochemical changes in the prefrontal cortex (PFC) of cocaine-dependent subjects.5 In preclinical studies, VEN also attenuated the acquisition of heroin self-administration and morphine dependence among rats in the conditioned place preference (CPP) model.6,7 Interestingly, VEN (10 mg/kg) treatment for 21 days has been reported to block evoked glutamate release in rats, suggesting a possible role of the glutamatergic system in VEN’s action.8 Furthermore, the release of glutamate in brain regions responsible for mediating drug-seeking behaviors was reported to cause relapse to cocaine-seeking behaviors, and the release of glutamate is critical in the relapse to METH and other drug-seeking behaviors.9,10 Based on previous studies, METH exposure can induce changes in glutamate homeostasis.11 Treatment with the well-known glutamate transporter type 1 (GLT-1) upregulator, ceftriaxone (CEF), has been documented to block relapse to use several drugs, including METH.12,13 Although CEF is effective in restoring glutamate homeostasis and preventing relapse to drug-seeking behaviors, it still has some drawbacks. In particular, CEF is an antibiotic with poor brain penetrability and cannot be given orally. Venlafaxine, however, is a novel antidepressant that has a unique pharmacological profile. Unlike other antidepressants, VEN does not inhibit monoamine oxidase (MAO) and does not change the activity of other receptors, such as adrenoreceptors, histaminergic receptors, or muscarinic receptors.14,15 Thus, it has fewer side effects when compared to other non-specific antidepressants. Furthermore, unlike CEF, VEN is not an antibiotic and can be given orally. Therefore, VEN is a strong candidate to be tested in the prevention of relapse to METH-seeking behavior.
Venlafaxine treatment for 21 days is hypothesized to prevent the reinstatement of METH-induced CPP in rats. It has been reported that VEN, like other antidepressants, needs chronic use to induce effective neurochemical changes and to produce an effect. For this reason, VEN was administered for 21 days in this study, the same dosing paradigm documented to block evoked glutamate release, as previously shown.8
Methods
Animals
Male Wistar rats (body weight per rat at the start of the experiments = 200-300 g) were provided by the Saudi Food and Drug Authority (SFDA; Riyadh, Saudi Arabia). The rats were held in regular plastic cages in a controlled environment with a 12:12 light–dark cycle, humidity = 30%, and temperature = 21°C. The animals had ad libitum intake of food and water. The experimental procedures performed on the animals were approved by the Research Ethics Committee at Taif University in accordance with the guidelines of the Institutional Animal Care and Use Committee of the National Institutes of Health and the Guide for the Care and Use of Laboratory Animals (Institute of Laboratory Animal Resources, Commission on Life Sciences, 1.96).
Drugs
Methamphetamine was obtained from the General Directorate of Narcotics Control in Saudi Arabia. Venlafaxine was provided by Jamjoom Pharmaceuticals (Jeddah, SA). Sterile saline solution (0.9% NaCl) was used to dissolve all drugs in this study.
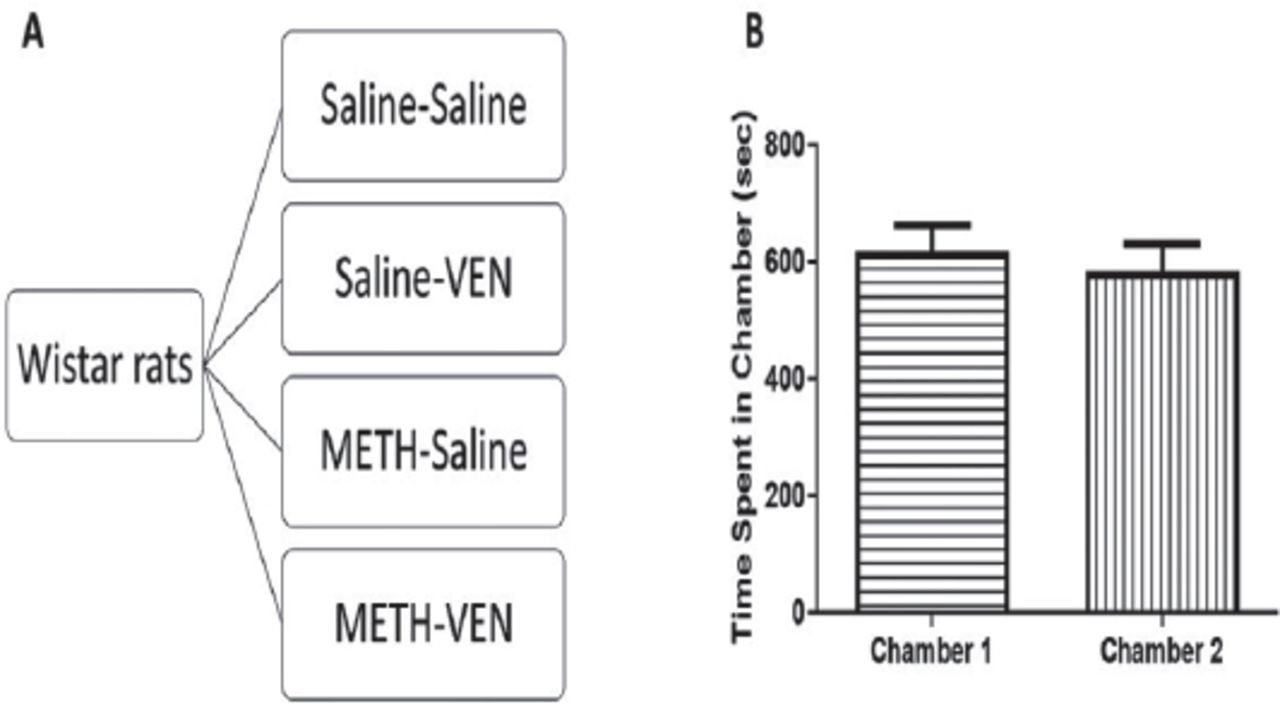
The experimental schedule and design are illustrated in Figures 1 & 2A. Following the acquisition of METH CPP and during extinction training, animals were randomly divided to receive saline or VEN instead of METH. The reinstatement was initiated by re-exposure to a single dose of METH to test any anti-reinstatement effects of VEN. Animals were assigned into 4 groups, as follows: a) Saline-Saline: rats (n=8) in this control group received saline during all phases of CPP. b) Saline-VEN: rats (n=8) in this group received saline during the conditioning phase and VEN (10 mg/kg, i.p.) during the extinction phase, and saline again during reinstatement. c) METH-Saline: during the conditioning phase, rats (n=7) in this group received METH (1 mg/kg, i.p.), saline during the extinction, and re-exposed to METH (1 mg/kg, i.p.) during the reinstatement phase. d) METH-VEN in which METH (1 mg/kg, i.p.) was administered to rats (n=7) in the conditioning phase, VEN (10 mg/kg, i.p.) was administered during the extinction phase, and METH (1 mg/kg, i.p.) was administered in the reinstatement phase.
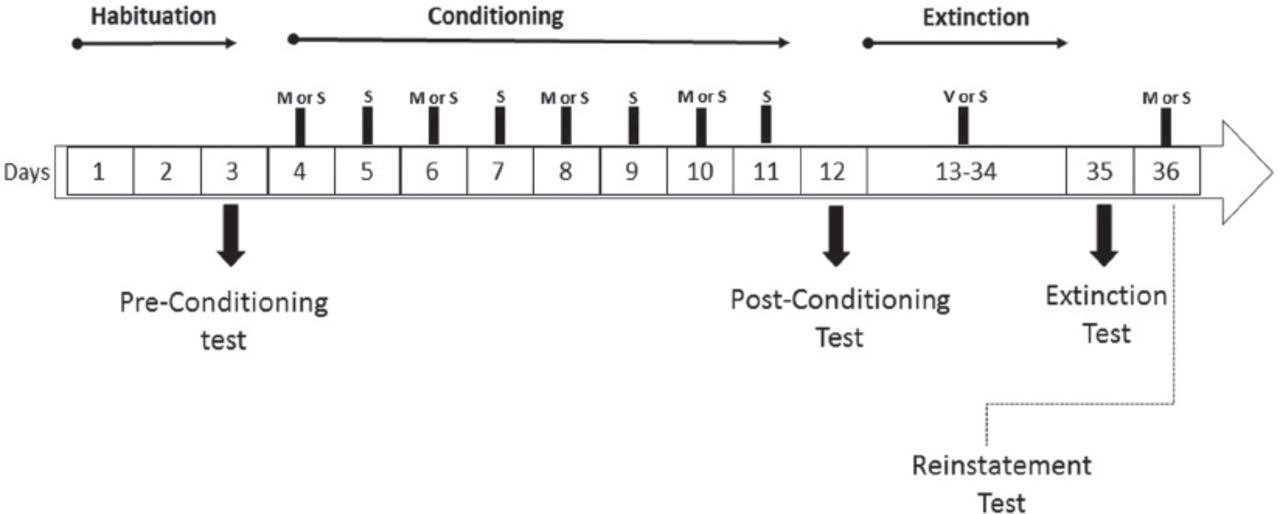
Experimental schedule of the conditioned place preference. M - methamphetamine, S - saline, V - Venlafaxine
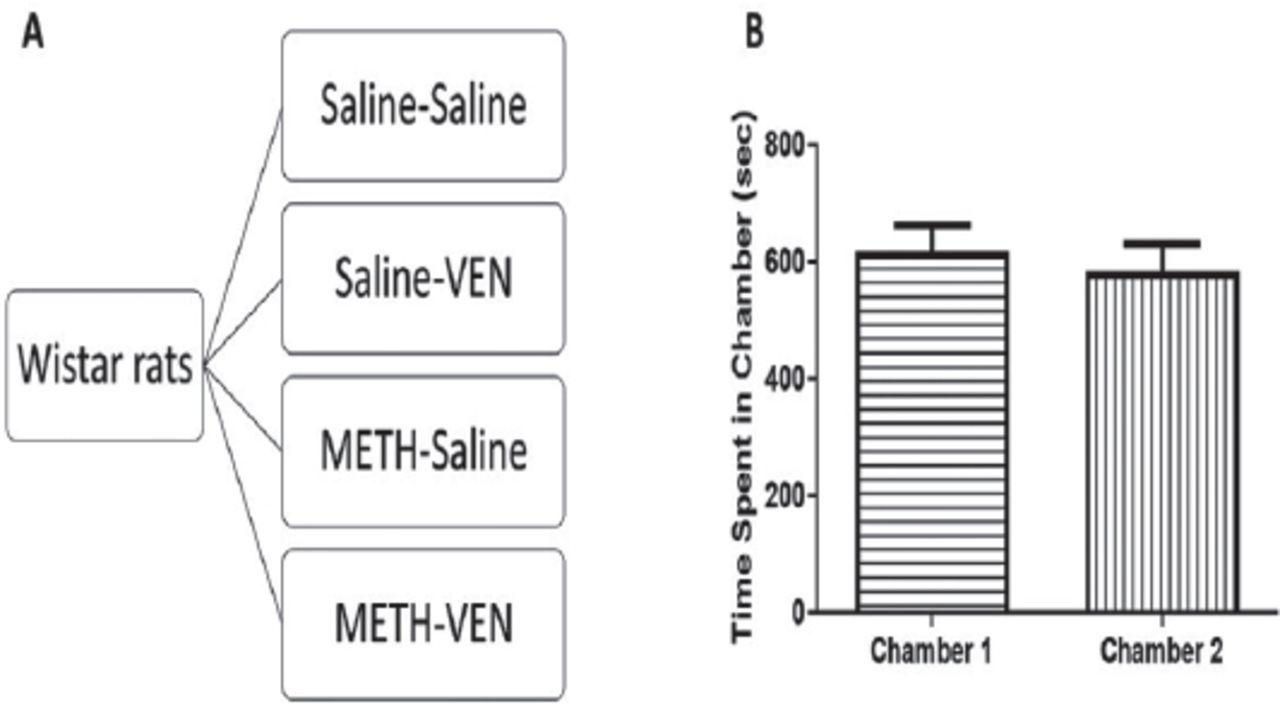
Experimental design A) Wistar rats were randomly assigned into four groups: 1) Saline-Saline-animals received saline during the conditioning, extinction, and reinstatement phases; 2) Saline-VEN-animals received saline in the conditioning, VEN (10 mg/kg, i.p.) in the extinction, and saline in the reinstatement phase; 3) METH-Saline, rats received METH (1 mg/kg, i.p.) in the conditioning, saline in the extinction, and METH (1 mg/kg, i.p.) in the reinstatement phase; 4) METH-VEN group in which METH (1 mg/kg, i.p.) was administered in the conditioning phase, VEN (10 mg/kg, i.p.) during the extinction phase, and METH (1 mg/kg, i.p.) was administered in the reinstatement phase. Time spent in each chamber during pre-conditioning and before any treatment. B) No significant changes in time spent were found in chamber 1 and chamber 2 during the preconditioning phase. Values shown as means ± S.E.M.
Venlafaxine (10 mg/kg) was administered intraperitoneally for 21 days which is the same dosing paradigm documented to block evoked glutamate release, as previously shown.8
Conditioned place preference paradigm
An acrylic CPP apparatus was designed to have 2 identically sized conditioning chambers (35 cm × 35 cm × 50 cm). Both visual and tactile cues distinguished the 2 conditioning chambers. The first chamber had horizontal black-and-white stripes with textured walls (Chamber 1). The inner walls of Chamber 2 were smooth, with vertical stripes. The floor of Chamber 1 was perforated with round holes. The floor of Chamber 2 was perforated with rectangular holes.
Pre-conditioning phase
Day 1 was considered the first day of the preconditioning training. On Days 1-3, each rat was allowed to explore both chambers for 20 min. On Day 3, this exploration was recorded by a digital camera (pre-conditioning test). An ANY-maze video tracking system was used to count the time the rats spent on each conditioning chamber. Rats spending more than 65% of their time in one chamber were considered to have a strong initial preference and were removed from the study, as previously shown.16,17
Conditioning phase
During the conditioning phase (Days 4-11), each rat received METH (1 mg/kg, i.p.) and was placed in the corresponding chamber with the door closed for 20 minutes session. For randomization purposes, 50% of rats received METH and placed in chamber 1 and the remaining received METH and placed in chamber 2. On the following day, each rat received saline i.p. and was placed in the other chamber with the door closed for 20 minutes. The process was repeated until 8 conditioning sessions were completed
On Day 12, each rat was allowed to explore both chambers for 20 minutes (post-conditioning test).
Extinction phase
Starting on Day 13, each rat in the experimental group received VEN (10 mg/kg, i.p.) while rats in the control groups received saline in their home cages. Treatment with either VEN or saline was given daily for 21 days. During this treatment period, each rat had free access to both chambers for 20 minutes per day. On Day 35, each rat was allowed to explore both chambers for 20 minutes (post-extinction test). For rats to be considered extinguished and to complete the study, time spent in the post-extinction test compared to the post-conditioning test should be reduced by 25%, as described previously.13
Reinstatement phase
On Day 36, rats in the experimental groups were re-exposed to METH (1 mg/kg) and allowed to explore the CPP apparatus for 20 minutes (reinstatement test).
Locomotor activity
Any possible sedative effects of VEN that might affect the reinstatement of METH CPP were investigated by assessing motor activity. Motor activity was assessed during the preconditioning, conditioning, extinction, and reinstatement phases by quantifying total distance traveled, using an ANY-maze video tracking system.
Statistical analysis
Two-way repeated measure ANOVA (Time x Chamber) was used to analyze time spent in conditioning chambers. When significant main effects or interactions were found, Newman-Keuls multiple comparisons were performed. Distance traveled was analyzed using one-way ANOVA, followed by the Newman-Keuls multiple comparisons test. All data were statistically analyzed by GraphPad Prism, using a 0.05 level of significance.
Results
Initial preference for each chamber during the pre-conditioning phase
The habituation sessions were conducted over 3 days. On the third day, the time spent in each chamber was calculated, which is considered the pre-conditioning results or initial preference for each chamber. The paired 2-tailed t-test revealed no significant difference between time spent in Chamber 1 compared to Chamber 2 (t=0.3503, df=30, p=0.7285; Figure 2B).
Effect of VEN on the reinstatement of METH CPP
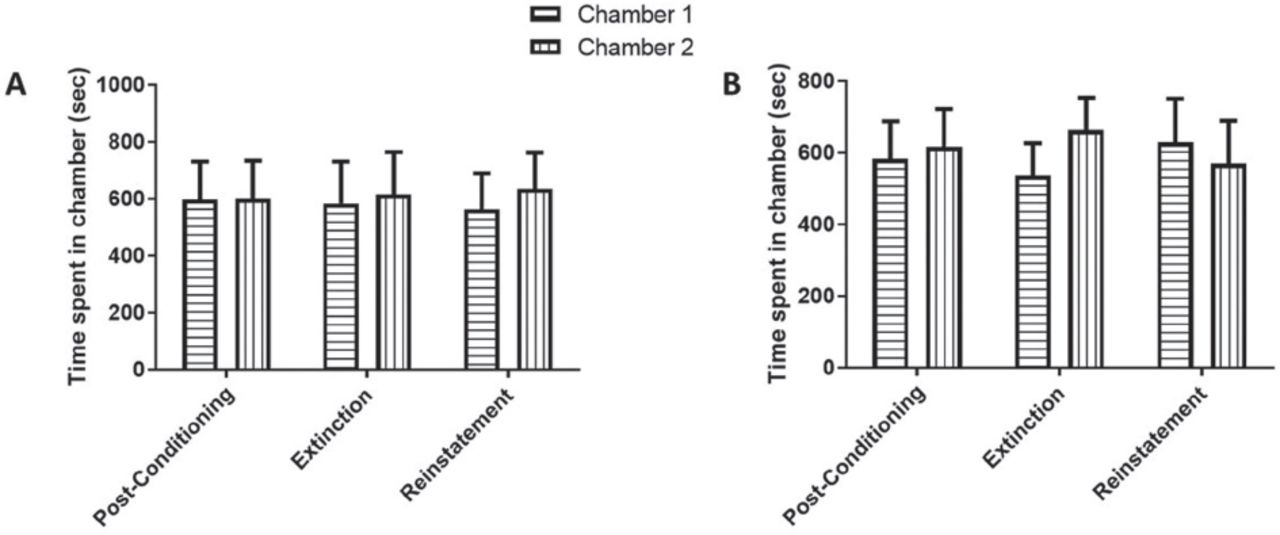
In Experiment 1, we tested the effect of administering only saline on the preference tests during post-conditioning, extinction, and reinstatement, to investigate any effects of saline alone on our CPP paradigm. Saline was administered during both the conditioning and extinction phases. Finally, animals were injected with a single dose of saline and tested for CPP. No significant effect of time [F (2, 14) = 1, p=0.3927], non-significant effect of chamber [F (1, 7) = 0.0182, p=0.8965], and no significant interaction between time and chamber [F (2, 14) = 0.5183, p<0.6065] were revealed by using 2-way repeated measures ANOVA (Figure 3A).
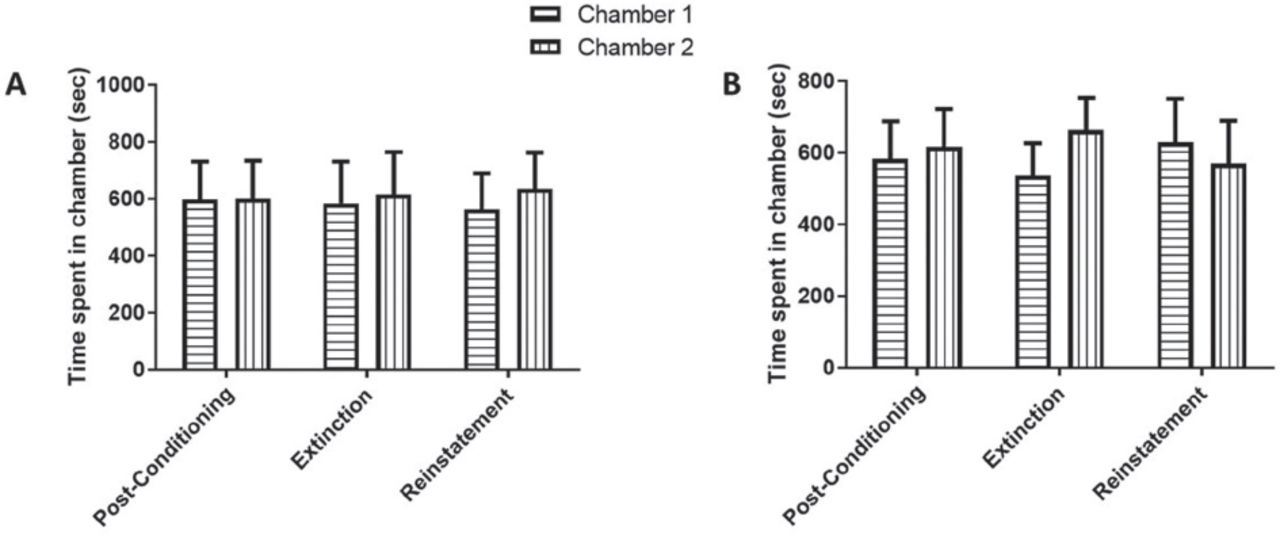
Time spent in the conditioning chambers during phases of post-conditioning, extinction, and reinstatement in A) Saline-Saline and B) Saline-VEN treated groups. No significant changes in time spent were found in Chamber 1 and Chamber 2 during all tested phases. Values shown as means ± S.E.M.
In Experiment 2, we tested the effect of administering saline on the preference tests during the post-conditioning phase and VEN during the extinction phase, to investigate any effects of VEN on our CPP paradigm. Saline was administered during the conditioning phase and VEN during the extinction phase. Animals were re-exposed to a single dose of saline and tested for CPP. No significant effect of time [F (2, 14) = 0.4667, p=0.6365], non-significant effect of chamber [F (1, 7) = 0.04135, p=0.8446], and no significant interaction between time and chamber [F (2, 14) = 0.3092, p=0.7389] were revealed by using 2-way repeated measures ANOVA (Figure 3B).
In Experiment 3, we tested the effect of METH on the CPP paradigm. Methamphetamine was administered during the conditioning phase, and saline was administered during the extinction phase. Animals were re-exposed to a single dose of METH and tested for CPP. A non-significant effect of time [F (2, 12) = 1, p=0.3966], significant effect of chamber [F (1, 6) = 6.941, p=0.0388], and a significant interaction between time and chamber [F (2, 12) = 20.45, p=0.0001] were shown by using 2-way repeated measures ANOVA. By using the Newman-Keuls multiple comparisons test, a significant increase in time spent was revealed during the post-conditioning test in the METH-paired chamber as compared to the other chamber (p<0.0001; Figure 4A). Time spent in the METH-paired chamber was significantly decreased in the extinction test compared to the post-conditioning test (p<0.01; Figure 4A).

Time spent in the conditioning chambers during phases of post-conditioning, extinction, and reinstatement in A) METH-Saline and B) METH-VEN. In METH-Saline group, rats spent time in the METH-paired chamber that was significantly elevated during the reinstatement test compared to the extinction test. VEN blocked reinstatement induced by re-exposure to METH. Values shown as means ± S.E.M. ***p<0.001 and ****p<0.0001. (@p<0.05 and @@p<0.01 compared to post-conditioning, while $$p<0.01 as compared to the extinction test).
Re-exposure to a single dose of METH significantly increased the time spent in the METH-paired chamber compared to the other chamber (p<0.0001). The time spent in the METH-paired chamber was significantly increased in the reinstatement test as compared to the extinction test (p<0.01).
In Experiment 4, we tested the effect of VEN on the reinstatement of METH in the CPP paradigm. Methamphetamine was administered during the conditioning phase and VEN during the extinction phase. Animals were re-exposed to a single dose of METH and tested for CPP. A non-significant effect of time [F (2, 12) = 1, p=0.3966], non-significant effect of chamber [F (1, 6) = 3.043, p=0.1317], and a significant interaction between time and chamber [F (2, 12) = 13.94, p=0.0007] were revealed by using 2-way repeated measures ANOVA revealed. A significant increase in time spent was revealed during the post-conditioning test in the METH-paired as compared to the other chamber (p<0.001; Figure 4B). Time spent in the METH-paired chamber was significantly decreased in the extinction test compared to the post-conditioning test (p<0.05; Figure 4B).
Re-exposure to a single dose of METH did not significantly change the time spent in the METH-paired chamber when compared to the other chamber (p>0.05). The time spent in the METH-paired chamber was not significantly changed in the reinstatement test as compared to the extinction test (p>0.05).
Effect of VEN and METH on locomotor activity
We tested the distance traveled during the preconditioning phase in the Saline-Saline, Saline-VEN, METH-Saline, and METH-VEN groups. One-way ANOVA revealed no significant changes in distance traveled among the Saline-Saline, Saline-VEN, METH-Saline, and METH-VEN groups during either the preconditioning phase [F (3, 28) = 0.8095, p=0.4993, Figure 5A], conditioning phase [F (3, 28) = 1.39, p=0.2663, Figure 5B], or extinction phase [F (3, 28) = 0.8095, p=0.4993; Figure 5C]. However, during the reinstatement phase, one-way ANOVA revealed a significant main effect among the Saline-Saline, Saline-VEN, METH-Saline, and METH-VEN groups [F (3, 26) = 3.334, p=0.0347; Figure 5D]. The Newman-Keuls multiple comparisons test showed a significant increase in distance traveled in the METH-Saline and METH-VEN treated groups compared to its corresponding control groups (p<0.05; Figure 5D).
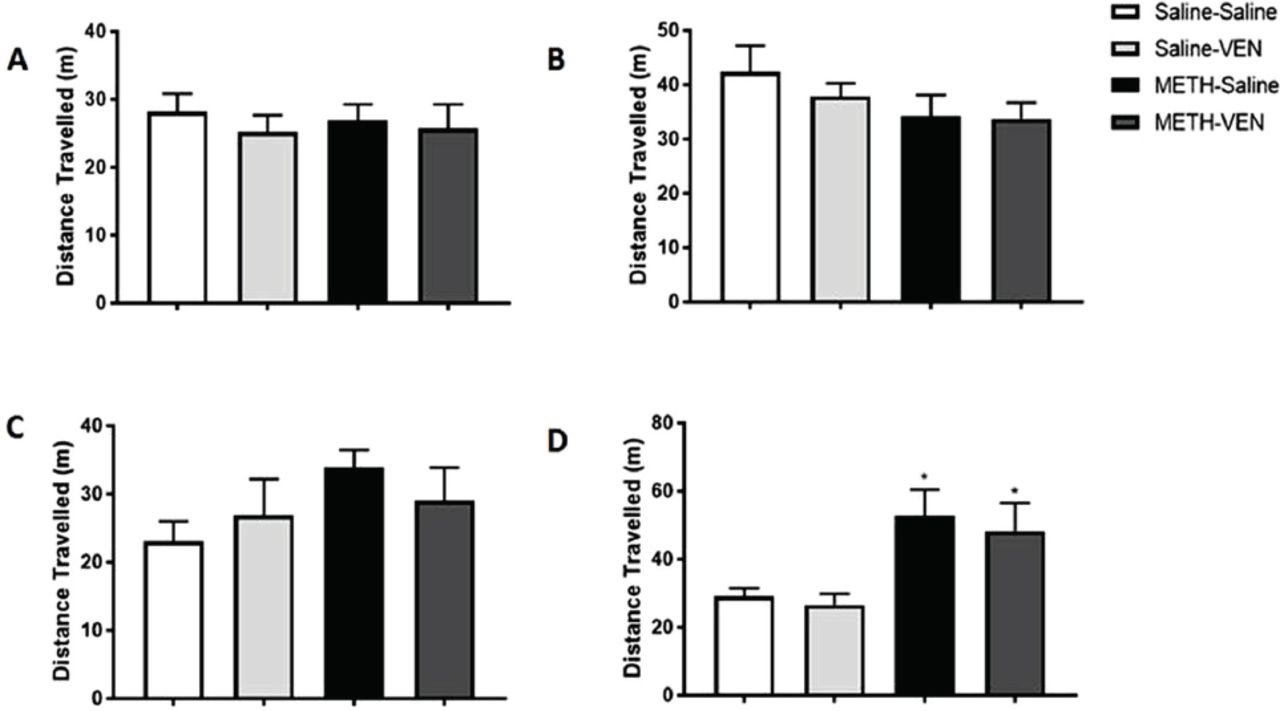
Distance traveled during Pre-conditioning (A), Conditioning (B), Extinction (C), and Reinstatement (D) phases in Saline-Saline, Saline-VEN, METH-Saline, and METH-VEN treated groups. During the reinstatement phase (D), distance traveled was significantly increased in the METH-Saline and METH-VEN treated groups compared to the corresponding control groups. Distance traveled was not significantly changed among all groups during Pre-conditioning (A), Conditioning (B), and Extinction (C) phases. Values shown as means ± S.E.M. *p<0.05.
Discussion
In this study, VEN treatment has been shown, for the first time, to inhibit METH-induced reinstatement of CPP in a rat model of relapse to drug use. This effect of VEN is in accordance with previous studies that showed the beneficial effects of VEN in attenuating the acquisition of heroin self-administration and morphine dependence in the CPP model in rats.6,7 The anti-reinstatement effects of VEN cannot be attributed to non-specific effects on locomotor behavior since VEN did not affect distance traveled in this study. This beneficial effect of VEN on relapse to METH CPP could be due to its antidepressant effects. This is in line with a previous report showing that chronic, but not acute, treatment with VEN blocked opiate seeking,7 suggesting the involvement of antidepressant effects. Importantly, abstinence from METH has been shown to be associated with symptoms of depression, such as anhedonia, loss of concentration, and irritability in humans.18 Moreover, stress is one of the main triggers of relapse to drug seeking and also one of the main features and causes of depression.19 Of note, subjects with a previous history of substance use have been shown to be more prone to depression and vice versa.20 In this study, VEN antidepressant effects might alleviate depression associated with METH abstinence during the extinction phase. This might explain, in part, the beneficial effects of VEN found here in preventing relapse to METH seeking.
Another possible mechanism is that VEN and other antidepressants have been reported to block evoked glutamate release, suggesting a possible role of the glutamatergic system.8,21 Of note, glutamate release, during exposure to different triggers of relapse, has been shown to mediate relapse to drug-seeking behavior.22 Glutamate is mainly transported into astrocytes by the action of glutamate transporters.23 Astrocytic glutamate is then released into extrasynaptic space in exchange for extracellular cystine. This extrasynaptic glutamate can then activate presynaptic type II metabotropic glutamate receptors (mGluR2/3), which causes feedback inhibition of the release of glutamate.24 Augmented glutamate release due to decreased glutamatergic tone on mGluR2/3 has been shown to occur during exposure to different triggers of relapse to drug seeking.25 Treatment with LY379268, a mGluR2/3 agonist, has been reported to block relapse to heroin, ethanol, METH, and cocaine seeking during exposure to different triggers of relapse, such as context-, stress-, drug-, and cue-induced relapse to drug seeking.10,26-29 Thus, blocking glutamate release during exposure to different triggers of relapse should be an effective strategy in preventing relapse to drug seeking. Therefore, the effects of VEN in preventing relapse to METH seeking might be explained by blocking the release of glutamate during reinstatement. Further studies are needed to explore the effects of VEN on glutamate transporters expression and homeostasis.
In the management of drug addiction, relapse prevention for drug use is the essential goal. Unfortunately, most treatment approaches for METH dependence were not promising in clinical trials, and there is no approved treatment for this disorder.30-33 However, some promising findings of modafinil, a wake-promoting drug, in the management of METH addiction have been shown in some clinical trials.34,35 In preclinical studies, modafinil treatment prevented the relapse to METH self-administration in rats.36 Modafinil also blocked relapse to morphine CPP by a possible involvement of the glutamatergic system in its action.37 Furthermore, treatment using the well-established GLT-1 upregulator, CEF, was successful in preventing relapse to METH CPP in rats.13 Although modafinil and CEF were both found useful in blocking relapse to METH seeking in animal models, these drugs have some limitations. Antibacterial effects, poor oral bioavailability, and limited brain penetrability are important limitations of CEF. The abuse liability of modafinil, especially in case of a previous history of stimulant dependence, has caused this drug to be labelled as a controlled medication along with its cardiovascular adverse effects.38-40 For these reasons, VEN might be a more effective and better drug candidate in preventing relapse to METH seeking compared to modafinil and CEF.
In conclusion, a reinstatement of METH CPP was established by a single dose of METH in extinguished rats. Venlafaxine treatment for 21 days prevented the reinstatement of METH CPP. The anti-reinstatement effect of VEN might be mediated by its antidepressant action as well as its effects on the glutamatergic system. These findings suggest a possible drug candidate in the management of METH dependence.
Acknowledgment
The work was supported by Research grant 1-438-5836 from Deanship of Scientific Research, Taif University, SA. The author thanks the General Directorate of Narcotics Control in Saudi Arabia for their cooperation in providing samples of METH. The author would like to thank Atiah Almalki, Omar Alzahrani, Hussam Almalki, and Abdulrahman Nasr for their help in the experiments. The author thanks the Saudi Food and Drug Authority for generously providing the animals. The author also thanks Jamjoom Pharmaceuticals for providing VEN.
Footnotes
Disclosure. The author have no conflict of interests, and the work was not supported or funded by any drug company. The work was supported by the Deanship of Scientific Research, Taif University, Saudi Arabia (Research Grant 1-438-5836).
- Received December 19, 2018.
- Accepted March 10, 2019.
- Copyright: © Saudi Medical Journal
This is an open-access article distributed under the terms of the Creative Commons Attribution-Noncommercial-Share Alike 3.0 Unported, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.