


Abstract
Objectives: To investigate the effect of androgens and estrogens on surtuin 1 (SIRT1) expression in human aortic endothelial cells (HAECs).
Methods: Real-time polymerase chain reaction analysis of SIRT-1 expression over 48 hours (h) was performed in HAECs treated with various concentrations of dehydroepiandrostendione (DHEA), androstenedione and testosterone (androgens), estrone (E1), estradiol (E2), and estriol (E3) (estrogens) to investigate the dose-dependency of time courses. The influence of high glucose on SIRT1 expression induced by the androgens and estrogens was also examined.
Results: Dehydroepiandrostendione, androstenedione, and testosterone remarkably produced a dose-dependent increase in SIRT1 expression in the range of 10 to 20 μg/ml. High glucose (40mM) medium had significantly inhibitory effects on 10 μg/ml DHEA-induced SIRT1 expression (p=0.024). Estrone and E2, but not E3, caused a marked dose-dependent increase in SIRT1 expression from 10 to 20 μg/ml. Treatment with 20 mM or 40 mM glucose medium did not significantly inhibit E1- and E3-induced SIRT1 expression in control medium; however, both high glucose mediums significantly emphasized E2-induced SIRT1 expression in control medium (p=0.007, p=0.005).
Conclusion: These results suggest that DHEA, androstenedione, testosterone, E1, and E2 definitely activate SIRT1 expression in HAECs. A high glucose medium is potent to inhibit the basal gene expression; however, it could not reduce powerful androgen- and estrogen-induced SIRT1 expression in HAECs.
Delineating hormonal signaling changes that occur across a lifespan and searching interventions may improve the quality of life (QOL) of elderly people and activate longevity.1 Surtuin 1 (SIRT1) is a key gene associated with longevity. Surtuins regulate steroid hormone signaling through a variety of molecular mechanisms and modulate pathways that modify steroid hormone receptors through phosphorylation.2
Polyphenols and ω-3-line polyunsaturated fatty acids (PUFAs) contribute to anti-aging and are also closely associated with activation of SIRT1 expression in some cultured cells.3 Various hormones affect SIRT1 expression in these cells. Neurosteroids play an important role in cognitive function and QOL, and hormones such as androgens and estrogens are especially critical for prevention of atherosclerosis and ageing and promotion of longevity through activation of surtuins.4-7 Estrogen- and testosterone-dependent actions play a vital role in mitochondrial processes implicated in aging.8 Beneficial effects of dehydroepiandrostendione (DHEA), an androgen, as an anti-aging steroid have been shown in vitro and in vivo, including the stimulation of immunity and then the suppression of diabetes, atherosclerosis, dementia, obesity, and osteoporosis activities.9 Estrogens may also modulate cardiovascular health through expression of SIRT1, possibly in the AKT and ERK signal pathways.10 The beneficial effects of estradiol appear to occur from its antilipofuscin, antioxidant and anti-lipid peroxidation properties and to imply an overall anti-aging action.11 Selective targeting of estrogen receptor α (ERα) with an agonist potentiates chemotherapy for treatment of ovarian cancer, and suppression of SIRT1 may accelerate this therapeutic effect.12
A high glucose medium can inhibit the effect of many different agents. Testosterone regulates ß-cell mass, at least in part, by androgen receptor (AR) activation in ß-cells of male rats, and the ß-cell AR is degraded under hyperglycemic conditions.13 In contrast, Chakrabarti et al14 found that control of oxidative stress by adenosine monophosphate-activated protein kinase (AMPK) activation recovers normal estrogen responses, even in the presence of hyperglycemia. Furthermore, at high glucose, kaempferol targets estrogen-related receptor α and suppresses angiogenesis of human aortic endothelial cells (HAECs).15
Taken together, it has been known the sex hormones are potential to activate the gene associated with longevity and high glucose medium could abolish the hormones’ action; however, it is unknown regarding the topic in HAECs as human cells. In order to elucidate these prior unanswered questions, we examined the effects of androgens and estrogens on SIRT1 expression in HAECs, and we investigate whether a high glucose level can affect hormone-induced SIRT1 expression.
Methods
This cross sectional study was conducted at the Joint Laboratory Office (JLO), Dokkyo Medical University Saitama Medical Center, Saitama, Japan from April 2017 to March 2019. The present medical research does not involve human subjects and was approved by our institutional ethical review board, and it is not previously published in partial or full in the website or printed journal in other language than English.
Cell culture
Human aortic endothelial cells were gained from American Type Culture Collection (ATCC Manassas, VA, USA) and cultured in Medium200 containing 10% large vessel endothelial supplement (LVES) (Gibco, Grand Island, NY, USA) at 37°C in a 5% CO2 incubator (Thermo Fisher Scientific, Carlsbad, CA, USA), as similar as methods shown in our previous report.3 For researches of SIRT1 expression, cells were seeded (105 cell/ml) and cultured in medium with 10% LVES for 24 hours (h), with addition of vehicle (dimethyl sulfoxide [DMSO]), DHEA (Wako Pure Chemical Industries, Japan), androstenedione, testosterone, estrone, estradiol or estriol (Sigma-Aldrich, St. Louis, MO, USA). Cell culture was followed using normalization to DMSO-treated cells.
Assessment of cell viability
After each treatment, the numbers of total and live cells were calculated with trypan blue staining. Cell viability was estimated by counting the ratio of live cells using trypan blue exclusion.16
Quantitative real-time polymerase chain reaction (PCR) analysis
Total ribonucleic acid (RNA) was extracted from HAECs using an RNA assay Mini Kit (Qiagen, Hilden, Germany), as similar as methods shown in our previous report.3 The complementary deoxyribonucleic acid was compounded using a SuperScript3 first-strand synthesis system for reverse transcription polymerase chain reaction (RT-PCR) (Invitrogen Life Technologies, Carlsbad, CA, USA). The RT-PCR was conducted in duplicate using FastStart SYBR Green Master Super Mix (ROX) (Roche Applied Science, Basel, Switzerland) in a 7500 Fast RT-PCR System (Applied Biosystems, Foster City, CA, USA). Data were normalized to ß-actin and are presented as the SIRT1/ß-actin densitometric ratio.
Surtuin1 expression in HAECs was examined using real-time PCR analysis. Dehydroepiandrostendione, androstenedione and testosterone (androgens), and estrone (E1), estradiol (E2), and estriol (E3) (estrogens) were added to HAECs at various doses, and time courses of SIRT1 expression were determined. The effects of glucose (5 to 40 mM) on hormone-induced SIRT 1 gene expression were also examined.
Statistical analysis
Data are presented as the mean ± standard deviation (SD). Statistical analyses of means were performed by analysis of variance (ANOVA), followed by a Student t-test, using the Statistical Package for Social Sciences (Japan IBM, Tokyo, Japan). Comparisons among multiple groups were evaluated by Bonferroni test if the ANOVA was significant. A p<0.05 was considered statistically significant.
Results
Time course of SIRT1 expression
Surtuin1 expression at 6, 12, 24 and 48 h after treatment of HAECs was examined using quantitative RT-PCR analysis, with each experiment repeated 3 or 6 times. Surtuin1 expression was not observed at 6 and 12 h, but was detected at 24 and 48 h (data not shown). Therefore, SIRT1 expression at 24 h was used in all studies shown below.
Dose-dependence of SIRT1 expression
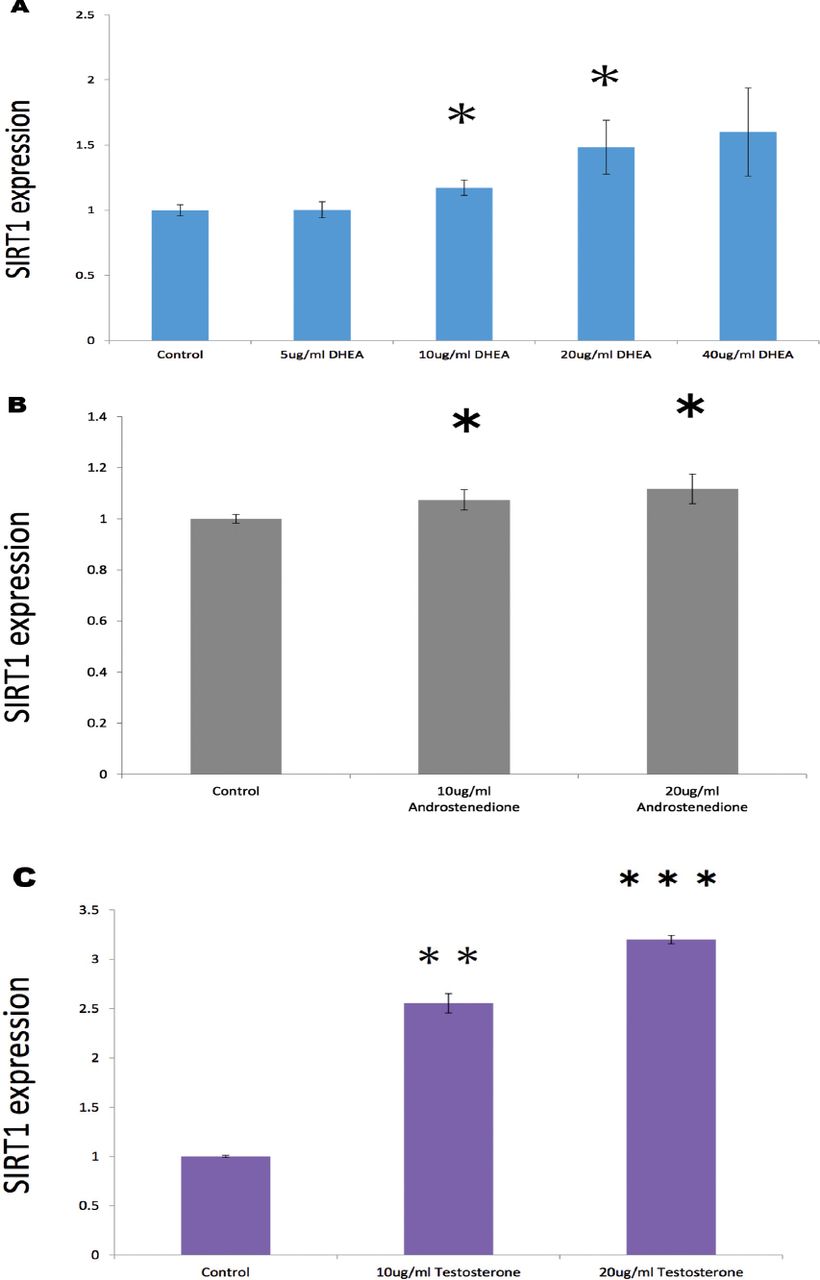
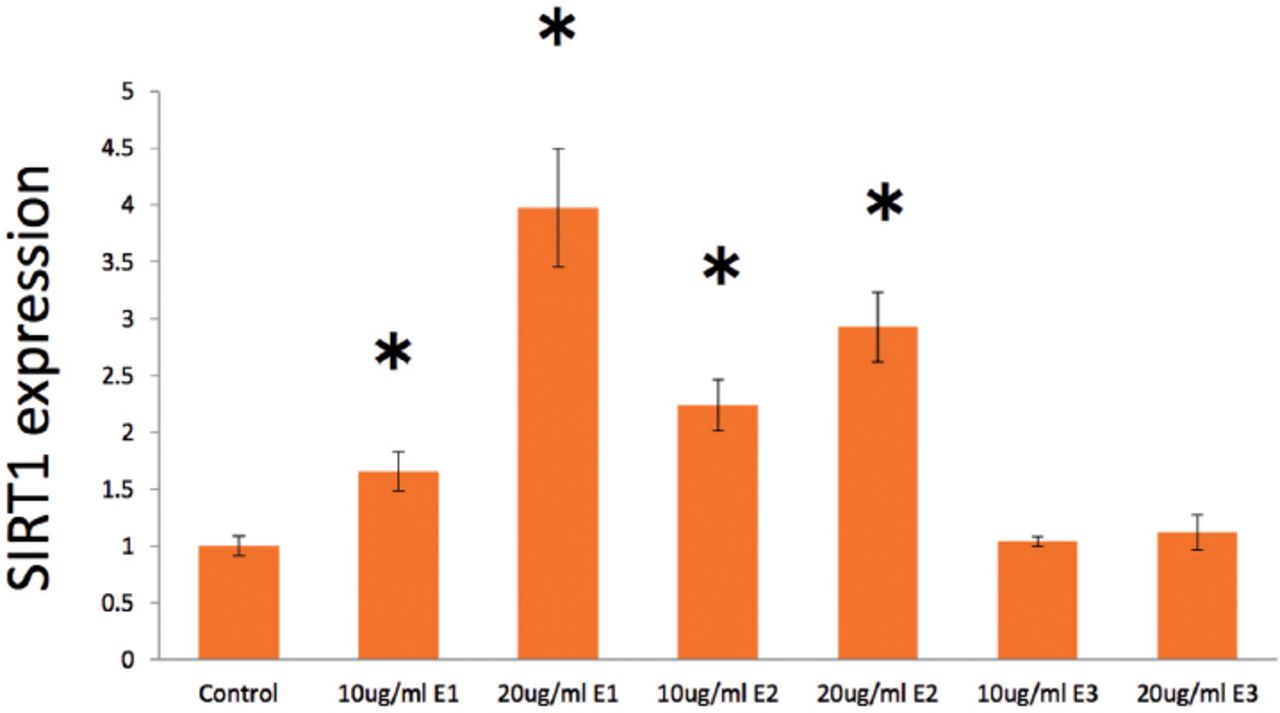
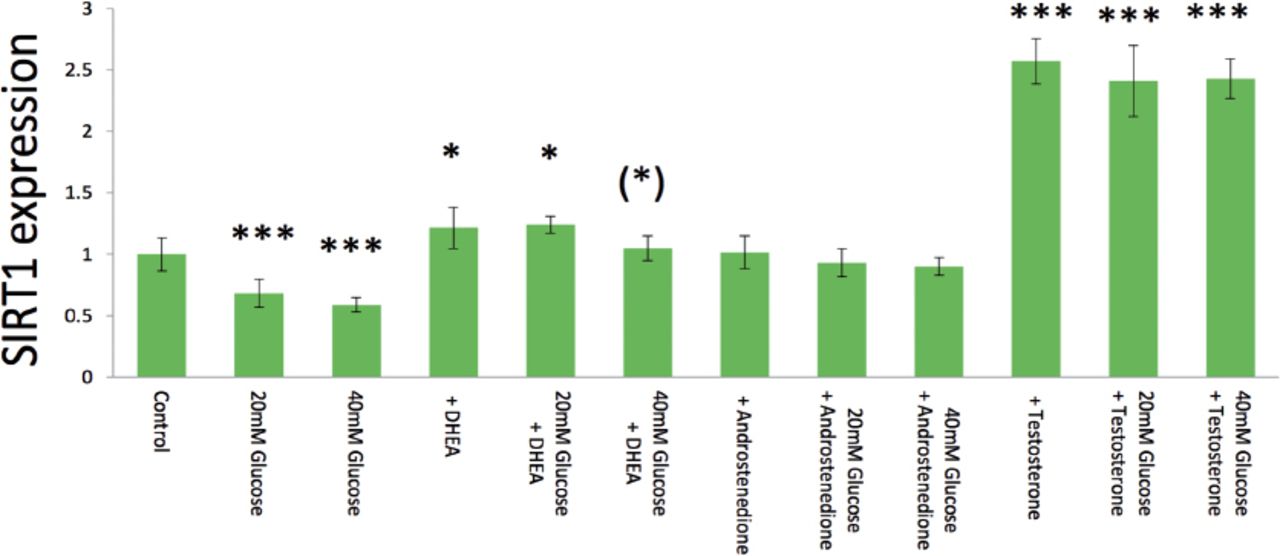
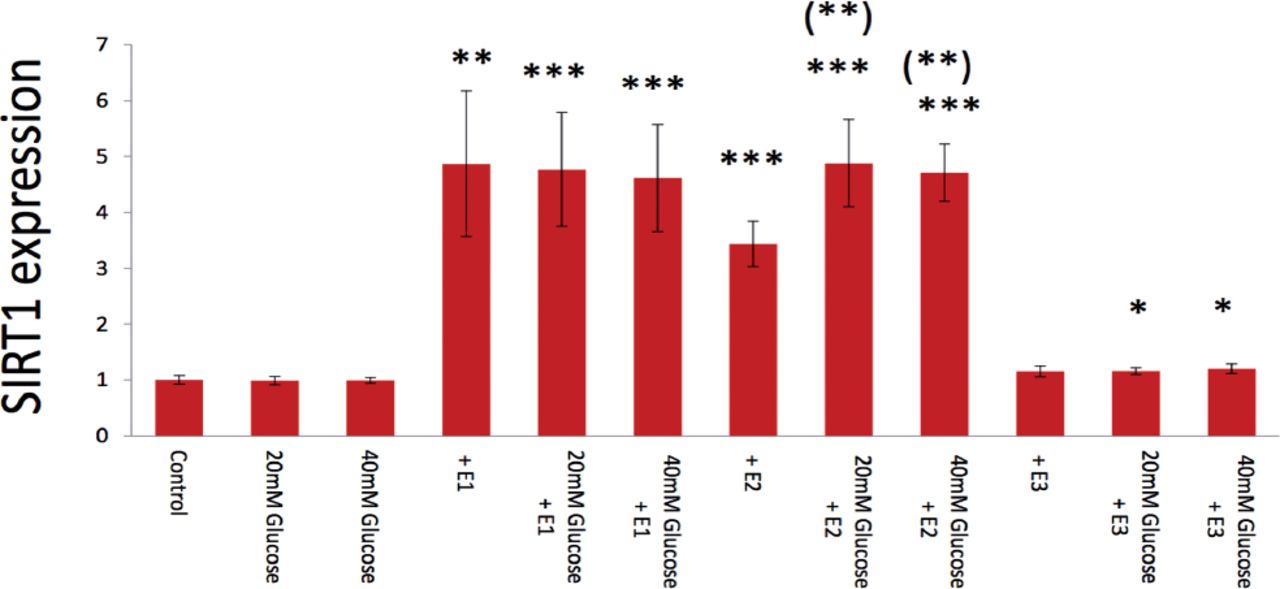
In treatment with androgens, SIRT1 expression was markedly and dose-dependently elevated by DHEA from 0 to 40 µg/ml (Figure 1A), especially both 10 µg/ml (p=0.013) and 20 µg/ml DHEA (p=0.027) were significant elevated compared with control values. Surtuin1 expression was also significantly affected by androstenedione from 10 to 20 µg/ml (Figure 1B), which showed that both 10 µg/ml androstenedione (p=0.020) and 20 µg/ml androstenedione (p=0.019) were significant stimulated on the gene expression. Furthermore, SIRT1 expression was dose-dependently elevated by testosterone in both 10 µg/ml (p=0.003) and 20 µ g/ml (p=0.0003) (Figure 1C). With estrogens, SIRT1 expression was significantly elevated dose-dependently by E1 from 10 (p=0.037) and 20 µg/ml (p=0.015) (Figure 2), with almost 4-times higher expression at 20 µg/ml E1 compared with control medium. Similarly, E2 from 10 (p=0.011) to 20 µ g/ml (p=0.010) dose-dependently increased SIRT1 expression with a significant alteration (Figure 2); however, E3 from 10 (p=0.314) to 20 µg/ml (p=0.261) had no significant effect on SIRT1 expression (Figure 2).

Effects of A) dehydroepiandrosterone (DHEA) (0-40 µ g/ml), B) androstenedione (0-20 µ g/ml), and C) testosterone (0-20 µ g/ml) on surtuin 1 (SIRT1) expression in human aortic endothelial cells at 24 hours after the start of treatment. Data are shown as the mean±SD of the SIRT1/ß-actin densitometric ratio with each agent relative to that with control medium determined in real-time polymerase chain reaction analysis in 2 separate experiments. *p<0.05, **p<0.01, ***p<0.001 versus control.
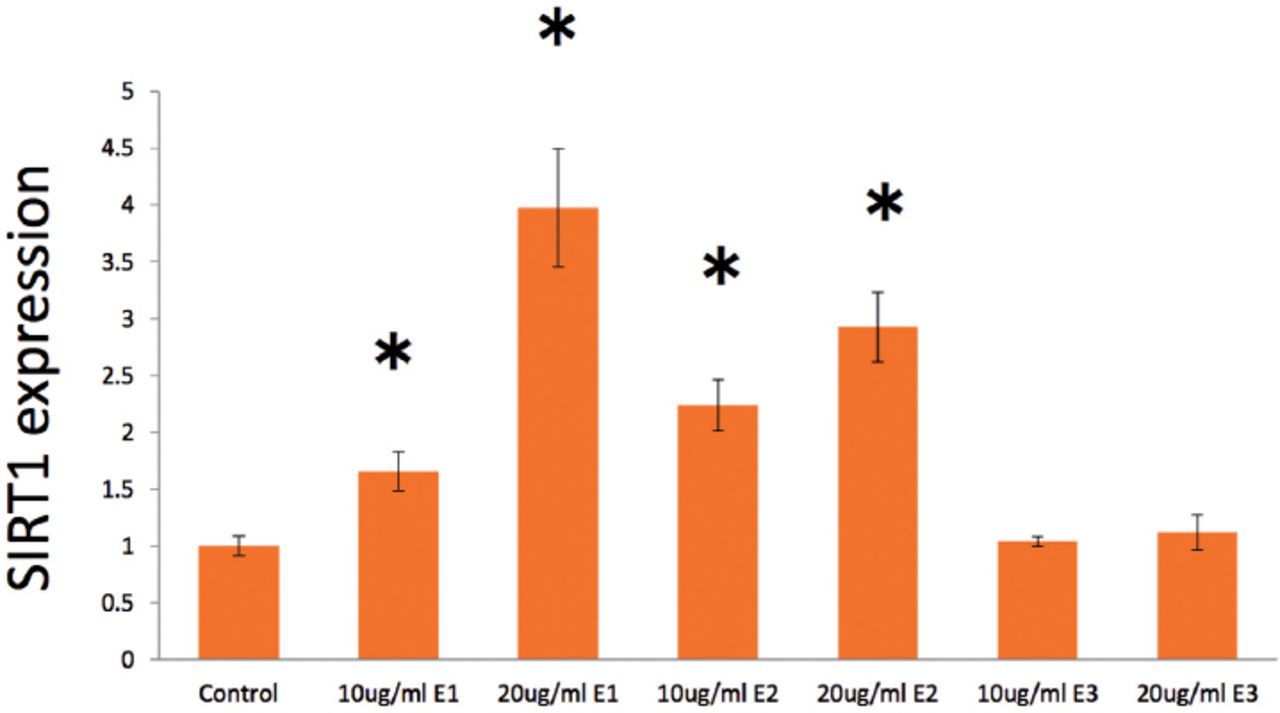
Effects of estrone, estradiol, and estriol (all 0-20µ g/ml) on surtuin 1 (SIRT1) expression in human aortic endothelial cells at 24 hours after the start of treatment. Data are shown as the mean ± SD of the SIRT1/ß-actin densitometric ratio with each agent relative to that with control medium determined in real-time polymerase chain reaction analysis in 2 separate experiments. *p<0.05 versus control.
Influence of high glucose on hormone-induced SIRT1 expression
Both 10 mM (p=0.0007) and 20 mM (p=0.0002) glucose significantly inhibited hormone-induced SIRT1 expression compared to control 5 mM glucose, and then 20 mM glucose tended to show greater inhibition than 10 mM glucose (Figure 3). With androgens, 40 mM glucose significantly inhibited 10 µg/ml DHEA-stimulated SIRT1 expression compared to 5 mM glucose did (p=0.024) (Figure 3). Both 20 mM (p=0.221) and 40 mM glucose (p=0.106) tended to inhibit SIRT1 expression induced by 10 µg/ml androstenedione with 5 mM glucose (control), but the differences were not statistically significant (Figure 3). As well as data from androstenedione, both 20 mM (p=0.205) and 40 mM glucose (p=0.098) tended to suppress 10 µg/ml testosterone -induced gene expression with 5 mM glucose (control); however, the differences did not lead to statistical significance (Figure 3).
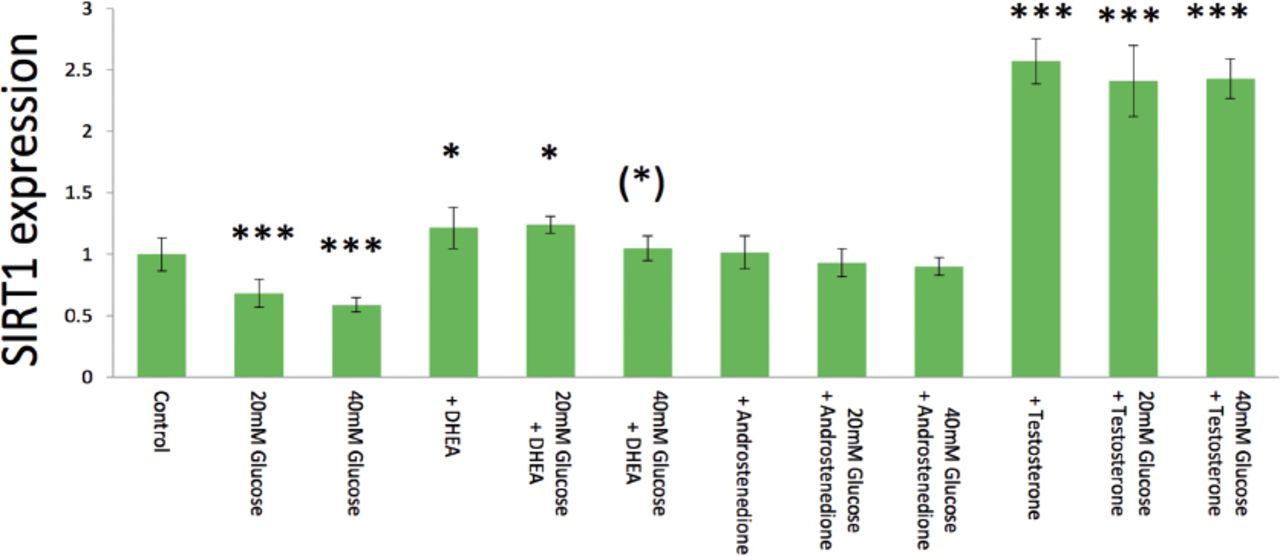
Glucose (20-40 mM) inhibition of basal, and dehydroepiandrosterone (DHEA), androstenedione-, and testosterone-induced surtuin 1 (SIRT1) expression in real-time polymerase chain reaction analysis. Data are shown as the mean±SD of the SIRT1/ß-actin densitometric ratio with each agent to that from control medium in 2 separate experiments. *p<0.05, ***p<0.001 versus (vs) control, and (*)p<0.05 vs. same agent-induced control medium
For estrogens, 20 mM (p=0.401) and 40 mM glucose (p=0.291) did not significantly inhibit SIRT1 expression stimulated by 10 µg/ml E1with 5mM glucose (control) (Figure 4). Similarly, 20 mM (p=0.465) and 40 mM glucose (p=0.302) did not show any significant inhibition on SIRT1 expression stimulated by 10 µg/ml E3 with 5mM glucose (control) (Figure 4). Interestingly, 20 mM (p=0.007) and 40 mM glucose (p=0.005) significantly emphasized SIRT1 expression induced by 10 µg/ml E2 with 5mM glucose (control) (Figure 4).

Glucose (20-40 mM) inhibition of E1-, E2-, and E3-induced SIRT1 expression in real-time PCR analysis. Data are shown as the mean±SD of the SIRT1/ß-actin densitometric ratio with each agent to that from control medium in 2 separate experiments. *p<0.05, **p<0.01, ***p<0.001 versus (vs) control, and (**) p<0.01 vs. same agent-induced control medium.
Discussion
The new and important aspects of the present study are that we successfully verified the activated effect of sex hormones such as DHEA, androstenedione and testosterone, E1, E2 on SIRT1 gene expression in HAECs, and then the inhibitory effect of high glucose medium on the gene expression, but not the inhibition of high glucose medium on those hormones-induced gene expression. We provided the novel information as regards the effect of sex hormones on SIRT1 gene expression in human culture cells such as HAECs through the present study, in addition to those effects already known in animal culture cells.
Decline of sex hormones is involved in the aging process and age-related diseases such as sarcopenia, fall, osteoporosis, cognitive decline, mood disorders, cardiovascular health, and sexual activity.17 Therefore, the actions of sex hormones on SIRT1 activity is of interest as a surrogate marker for longevity. In this study, we also investigated the influence of high glucose on these actions.
We found that the androgens DHEA and testosterone, but not androstenedione, augmented SIRT1 expression in a dose-dependent manner. These results are reasonable because DHEA and testosterone are more powerful bioactive hormones than androstenedione. A direct functional link with the AR is a critical determinant of progression of human prostate cancer and the sirtuins.5 Testosterone and SIRT have a critical role in prevention of vascular and neuronal aging.18 Based on our results, this effect of testosterone may be evoked through activation of the SIRT gene. Dehydroepiandrosterone inhibits primary rat Leydig cell proliferation by decreasing the cyclin mRNA level, and improves cells viability by modulating the permeability of the mitochondrial membrane and succinate dehydrogenase activity.19 These results might be due to the effect of DHEA on activation of SIRT expression in HAECs, as shown in the current study. Dehydroepiandrostendione prevents linoleic acid-induced endothelial senescence by restoring autophagy and autophagic flux through JNK activation, and the effect of DHEA on longevity might at least partly be associated with this mechanism in HAECs.20
We did not find an effect of androstenedione on SIRT1 expression, which is consistent with a report showing that androstenedione had no effect on development of bones, including sternebrae, and soft tissues.21 This suggests that androstenedione, a metabolite of the androgen pathway, might have weak bioactivity in cultured cells. Testosterone and SIRT1 have critical roles in prevention of vascular and neuronal aging, and the present results suggest that this may be partly due to testosterone activation of SIRT1 expression.18 Furthermore, testosterone upregulates the anti-aging klotho gene together with AR expression in the kidney, which supports our results despite the difference in the longevity gene.22 Novel metabolic pathways linked to male lifespan extension show that male-specific metabolomic responses to 17α-E2 depend on production of testicular hormones in adult life.23 This report and the present study show that testosterone plays an important role in longevity.
We also examined the effects of estrogens on SIRT expression. We found effects of E1 and E2, but not E3, on SIRT1 expression. Estrogens protect against ischemic stroke through prevention of neuron death via the SIRT1-dependent AMPK pathway, and this supports our results showing that E1 and E2 activate SIRT1 expression.24 Transvaginal E3 potentially offers a suitable physiologic delivery as a clinical benefit, however, E3 might be a weak activator of longevity, in contrast to E1 and E2.25 The anti-aging effect of 17ß-E2 is clear from lower lipid peroxidation and liver dysfunction parameters, as well as histological observation, but the mechanism of estrogen action on SIRT1 expression is unclear.26 The effects of estrogen and bisphenol A on expression of histone-modifying enzymes vary, and estrogen receptor and AR may both be important in therapeutic intervention.27 SIRT1 activities have also been linked to estrogenic effects through interaction with the G-protein coupled membrane bound estrogen receptor.28
There is widespread interest in sex hormones because of their apparent anti-aging effects. The Vitex agnus-castus fruit improves features of aging, such as oxidative stress, female sex hormone deficiency, and an atrophic endometrium.29 In general, anti-aging effects are associated with reducing oxidative stress and supplementation of female sex hormones. Thus, antioxidants (vitamin E, vitamin C, and carotenoids) and hormones (growth hormone, testosterone, DHEA, and vitamin D) can serve as anti-aging therapies.30 The present study focused on the bioactivities of sex hormones in anti-aging. A further study is needed to examine involvement of antioxidants on anti-aging in HAECs.
We found that high glucose had a significant suppressive effect on basal- and DHEA-induced SIRT1 expression in HAECs. We have also observed a similar phenomenon of inhibition of SIRT1 expression by high glucose in human monocyte leukemia cells, THP1 cells.3 High glucose may impair ERα transcriptional activity by inhibiting ß-catenin signaling in osteoblastic MC3T3-E1 cells, leading to decreased bone formation in a ligand-dependent or -independent manner.31 Testosterone prevents pancreatic ß-cell apoptosis due to glucotoxicity through reduction of expression of ATGR1 and its signaling pathway.32 In the present study, testosterone-induced SIRT1 expression was not inhibited by high glucose. Paradoxically, we observed augmentation of E2-induced SIRT1 expression by high glucose. Responsiveness to 17ß-E2 in primary human osteoblasts is modulated differentially by a high glucose concentration, which supports our results. Androgen replacement therapy could be effective for type 2 diabetes-induced ED and is a potential ED treatment,but we found that high glucose significantly inhibited DHEA-induced SIRT1 expression. Therefore, based on these previous reports and our results, androgen replacement therapy for T2DM-induced ED might be weakened by poor control of blood glucose.33,34
Activation of SIRT1 by resveratrol ameliorated cardiac injuries in a mouse model of dilated cardiomyopathy (DCM) through PGC-1 α-mediated mitochondrial regulation, and SIRT activation by sex hormones may be a therapeutic target for DCM.35 Control of oxidative stress by AMPK activation or antioxidants could restore normal estrogen responses, even in hyperglycemia.14 Consistent with this, we found that E1- and E3-induced SIRT1 expression was not diminished by high glucose. Testosterone regulates ß-cell mass, at least in part, by AR activation in ß-cells of male rats and the ß-cell AR is degraded under hyperglycemic conditions.13 Therefore, although testosterone-induced SIRT1 expression was not weakened under hyperglycemic conditions in vitro in the present study, hormone-induced SIRT1 expression might diminish in hyperglycemia in vivo. In contrast to high glucose, 2-deoxy-d-glucose (2-DG) extends the lifespan of Hs68 cells by increasing NAD+ levels and SIRT1 activity, and 2-DG has potential as a caloric restriction mimetic,therefore, 2-DG may activate SIRT1 expression in HAECs.
Study limitations
Although we applied 5 to 40 µ g/ml as the range of each agent’s concentration because of being worried about cell apoptosis due to drug toxicity, the range should be more wide to verify precisely those drugs’ effect. Furthermore, the present data were represented in RT-PCR analysis using each 3 wells in 2 separated experiments, respectively; however, the study should be performed using 4 wells in 3 separate experiments to avoid the variation of reappearance.
In conclusion, our results suggest that DHEA, androstenedione, testosterone, E1, and E2 activate SIRT1 expression in HAECs, and that high glucose can inhibit basal SIRT expression, but not that induced by androgens and estrogens. Future research is required to investigate whether other hormones besides sex hormones could activate SIRT1 gene expression in HAECs.
Acknowledgment
We are grateful to Prof. Daisuke Koya, Department of Diabetes and Endocrinology, Kanazawa Medical University, Ishikawa, Japan for his academic and technical advice. We also would like to thank PALABRA Inc. for English language editing.
Footnotes
Disclosure. This study was funded by the Department of Internal Medicine, Dokkyo Medical University Saitama Medical Center, Koshigaya, Japan.
- Received October 13, 2019.
- Accepted February 9, 2020.
- Copyright: © Saudi Medical Journal
This is an open-access article distributed under the terms of the Creative Commons Attribution-Noncommercial-Share Alike 3.0 Unported, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.